Introducción
Los bosques albergan
alrededor del 80% de la biodiversidad terrestre global, a pesar de ocupar
solamente el 31% de la superficie emergida (FAO 2020). Dentro de esta extensión forestal, aproximadamente la mitad se
mantiene en un estado relativamente intacto, de la cual, más de un tercio está
constituido por bosques primarios. En España, la superficie forestal abarca más
del 55% del territorio, con un total de 28 millones de hectáreas (ha), de las
cuales 18.5 millones son bosques (González Díaz et
al. 2020). De forma aún más concreta, el 56% se corresponde
a frondosas, 37% coníferas y un 7% a bosque mixto (González Díaz et
al. 2020). En Andalucía, a escala regional, predominan los
bosques mediterráneos del género Quercus (encina, alcornoque, quejigo y
roble melojo). Como ecosistema
forestal abierto en España y sobre todo en Andalucía, destacan las dehesas, que
representan sistemas silvopastoriles de bosque de dosel abierto donde se
mezclan árboles (del género Quercus principalmente en la zona
mediterránea) con pastizales u otras especies de matorrales (Ferraz-de-Oliveira et al. 2016). A escala nacional las dehesas cubren una superficie de 3.5 millones de
ha aproximadamente (7% del territorio español), de las cuáles 1.2 millones se
encuentran en la comunidad andaluza (14% del territorio andaluz) (Moreno y Pulido 2009). Por lo tanto, la degradación de estas áreas más
abiertas, por distintos motivos como el cambio climático y/o la pérdida de
biodiversidad, provocaría una pérdida irreversible de funciones y servicios
ecosistémicos (Marañón et al. 2012; Requena-Mullor et al. 2023). Por ejemplo, una disminución de los sumideros de carbono de los bosques
comprometería su papel como solución basada en la naturaleza para la mitigación
del cambio climático (Anderegg et al. 2020).
Entre los factores que
determinan la estructura de un bosque, definida como la manera en que la
vegetación está organizada en el espacio, teniendo en cuenta la variación y
dimensión de los individuos (Franklin et al. 2002), destaca el manejo humano (Franklin et al. 2002; Rendón-Pérez et al. 2021). Así, diferenciamos bosques sin manejo como aquéllos que carecen de
gestión formal, permitiendo un desarrollo natural del mismo (FAO 2003), de los bosques con manejo donde diferentes
técnicas de gestión, producción y/o conservación son aplicadas intensivamente (Hernández-Silva et al. 2018). Concretamente, el grado de intervención humana en los bosques de encina
o encinares (Quercus ilex) influye de manera significativa en su
estructura, funcionamiento y capacidad de resiliencia frente a eventos de
sequía. En bosques densos, donde la intervención es mínima, el sotobosque y la
cubierta forestal suelen ser más complejos, favoreciendo la retención de
humedad y la resiliencia frente a la escasez hídrica (Ellison et al. 2017). Sin embargo, estos bosques también pueden sufrir una mayor competencia
por recursos, lo que los hace más vulnerables durante sequías prolongadas. Por
otro lado, en los sistemas normalmente más manejados, como las dehesas, donde
se aclaran árboles y se combina el uso agrícola o ganadero, la estructura del
bosque cambia de forma drástica. Aunque la menor densidad de árboles permite
una mayor penetración de luz y reduce la competencia por agua, la capacidad de
estos ecosistemas para resistir la sequía puede verse comprometida por la
pérdida de la alta biodiversidad que albergan (Moreno et al. 2018; Parra-López et al. 2023), la disminución de la humedad edáfica y la simplificación del paisaje.
Los bosques intermedios, suelen tener un grado de manejo moderado, y
generalmente se sitúan en un punto medio en cuanto a resiliencia, ya que
presentan beneficios de ambos extremos, pero también riesgos asociados con la
alteración de su estructura natural (Pardos et al. 2021). En resumen, la intervención humana modifica el funcionamiento
ecológico del bosque mediterráneo, afectando tanto su capacidad de adaptación
al cambio climático como su estructura, lo que determina su respuesta a la
sequía y otros fenómenos climáticos extremos. Por ejemplo, el abandono del uso
tradicional para leña y carbón vegetal ha derivado en superficies de monte bajo
con altas espesuras, cuya competencia compromete el vigor y desarrollo de los árboles,
implicando una mayor vulnerabilidad a factores de estrés abióticos (sequía) o
bióticos (la seca, enfermedad causada por el hongo Phytophtora cinnamomi Rands.) (Mantero et al. 2020). También se ha incrementado el adehesamiento de los bosques densos
originales por la actividad humana, influyendo, además, en alteraciones de la
diversidad en sus componentes α, β y γ (Valladares et al.
2014). Otro factor a tener en cuenta es la aridez y
sequía intensificadas por el cambio climático, presentándose como factores
importantes de perturbación en los bosques mediterráneos andaluces, aumentando
el deterioro fisiológico de los árboles, los incendios y las enfermedades
forestales (Kim et al. 2017).
Comprender los procesos y dinámicas de los tipos de bosque,
en este caso de encina (Quercus ilex subsp. ballota),
proporcionará ventaja a la hora de anticipar, gestionar o mitigar, posibles
eventos futuros adversos que derivarán en estrategias de manejo más robustas.
Para afrontar estos desafíos de manera eficiente y rápida, es necesario hacer
uso de enfoques de modelación y monitoreo capaces de anticipar y guiar las
respuestas de los bosques al cambio global (López-Tirado y Hidalgo 2016). La combinación de datos medidos en campo, como
las parcelas de los inventarios forestales, junto a datos de teledetección
satelital, puede presentarse como una alternativa dinámica crucial ante datos
tradicionales más estáticos (Ruiz-Benito et al. 2017; Herraiz et al. 2025).
Entre otras ventajas, la teledetección permite un análisis retrospectivo de las
respuestas a la sequía, proporcionando información sobre los cambios temporales
en la productividad primaria del bosque y el contenido de agua del dosel en
grandes extensiones del territorio (Jiao et al. 2021). Entre
otros productos derivados de la teledetección, los índices espectrales de
vegetación (IVs) como el Índice de Vegetación de Diferencia Normalizada o NDVI
por sus siglas en inglés, así como el Índice de Salud de la Vegetación (VHI,
Vegetation Health Index), informan sobre su estado incluso durante eventos de
cambio climático extremos (Alcaraz-Segura et al. 2008; Bento et al. 2018; Vila-Viçosa et al. 2020).
En este sentido, si bien
la mayoría de los estudios sobre las respuestas de los ecosistemas forestales a
la sequía se han centrado en las respuestas de especies individuales (López-Tirado y Hidalgo 2016; Herraiz et al. 2023), es necesario conocer su respuesta, y resiliencia, a los eventos
extremos desde un punto de vista estructural y de gestión, y a nivel de paisaje
más amplio para periodos de tiempo más largos. Para ello, los bosques de encina
de Andalucía, definidos como densos (DEN), intermedios (INT) y abiertos (ABI)),
y con diferentes grados de gestión, constituyen un contexto geográfico idóneo
para testar nuestra investigación. Bajo estas premisas, en este trabajo
pretendemos mejorar la comprensión de las respuestas de los tipos de bosque de
encina en base a sus diferencias en estructura y composición de la vegetación y
características ambientales. De este modo, podemos analizar en un plazo amplio
de tiempo, las posibles y diferentes respuestas de cada tipo de bosque a
eventos adversos. De manera específica pretendemos: 1) conocer y examinar las
variables (bióticas, abióticas y espectrales) que caracterizan a cada tipo de
bosque de encina; 2) investigar las relaciones entre variables bióticas y
abióticas medidas en campo y los índices multiespectrales derivados de datos de
satélite; y 3) realizar un análisis multitemporal de los índices espectrales
para cada tipo de bosque y evaluar su relación con eventos climáticos
destacados. Las hipótesis que planteamos son: H1: existen
diferencias significativas entre los distintos tipos de bosque de encina
detectables con variables biofísicas medidas en campo e IVs obtenidos por
teledetección satelital; H2: Considerando que los IVs dependen
principalmente de factores climáticos (precipitación y temperatura) y
topográficos, la respuesta espectral de cada tipo de bosque estará influenciada
también por su estructura y gestión, reflejando una combinación única de
factores ambientales que influyen en ella; y H3: El análisis
interanual de los índices de vegetación permitirá observar una respuesta
diferente de cada tipo de bosque a eventos extremos producidos por el cambio
climático.
Materiales y métodos
Área y especie de estudio
El área de estudio abarca
toda la comunidad autónoma de Andalucía, situada en el sur de España con una
extensión de 87 609 km2. Andalucía está influenciada por la
región atlántica y mediterránea, predominando esta última, y presenta un rango
marcado de precipitación que coincide con un rango de sequía que se incrementa
de este a oeste de la comunidad. La temperatura media anual es suave (16 °C) y
con una elevada radiación solar (300 días soleados de media por año). En cuanto
a la elevación, Andalucía tiene un rango de 0 hasta 3500 m sobre el nivel del
mar (Sierra Nevada), con una elevación media de 1000 m. El relieve no sólo
ejerce efectos significativos en las condiciones climáticas, sino que también
influye notablemente en la vegetación presente y el tipo de ocupación del
suelo, dando lugar a una amplia variedad de tipos de coberturas vegetales con
distintos ciclos fenológicos y dinámicas (Caparros-Santiago y Rodríguez-Galiano 2020). Toda esta variabilidad a lo largo de la región
resulta en una gran variedad de especies, hábitats y ecosistemas. Tanto es así,
que hasta el 60% de todas las especies de flora y fauna existentes en la
península ibérica se pueden encontrar en Andalucía. Toda esta riqueza
medioambiental hace que más del 30% de la superficie de Andalucía presente
algún tipo de figura de protección para su conservación (Hernández et al. 2014).
En Andalucía hay una gran área ocupada por actividades
agrícolas de cultivos herbáceos y leñosos, destacando las áreas bajas del
Guadalquivir, así como depresiones entre montañas y las franjas costeras no
urbanizadas (Naranjo
Ramírez 2003). Por el contrario,
encontramos áreas en Sierra Morena y en las cordilleras Béticas donde
prevalecen las zonas boscosas con gran presencia de quercíneas y coníferas
respectivamente (Araque Jiménez y Sánchez Martínez 2009).
De entre todas las especies del género Quercus, la
encina (Quercus ilex subsp. ballota) es la más abundante en la
península ibérica y en Andalucía. Los encinares, por tanto, tienen una gran
importancia tanto funcional como de aportación de servicios ecosistémicos
(suministro, regulación, culturales) (Marañón et al. 2012; Kim et al. 2017; Bastias et al. 2025).
Por ende, se hace indispensable profundizar en el análisis y estudio de la
relación entre tipos de bosque de encina y las características del medio que
los rodea. No todos los bosques mediterráneos de encina presentan la misma
estructura y, por lo tanto, pueden tener dinámicas muy distintas a pesar de que
la especie dominante sea la misma. Pueden clasificarse de manera simplificada
en tres categorías en función de su densidad: abierto, intermedio y denso (Fig. 1). Los bosques con estructura abierta o
adehesados, se caracterizarían por presentar una densidad de árboles y
regeneración muy baja, y por consiguiente una biomasa escasa, aunque el
crecimiento relativo de los árboles podría ser mayor al haber menos competencia.
Además, se suelen encontrar en zonas llanas con baja elevación, facilitando la
intervención humana. Por el contrario, en el otro extremo tendríamos el bosque
denso, caracterizado normalmente por aparecer en zonas de elevada altura y
pendiente, con una cobertura de vegetación densa, y con escasa intervención
humana. Los bosques de tipo intermedio tendrían características intermedias
entre el abierto y el bosque denso (Fig. 1). Por
lo tanto, sería de gran utilidad poder distinguir qué características bióticas
y abióticas previamente relacionadas con datos de teledetección caracterizan
cada tipo de bosque para optimizar el desarrollo de estudios y/o trabajos de
conservación y monitoreo.
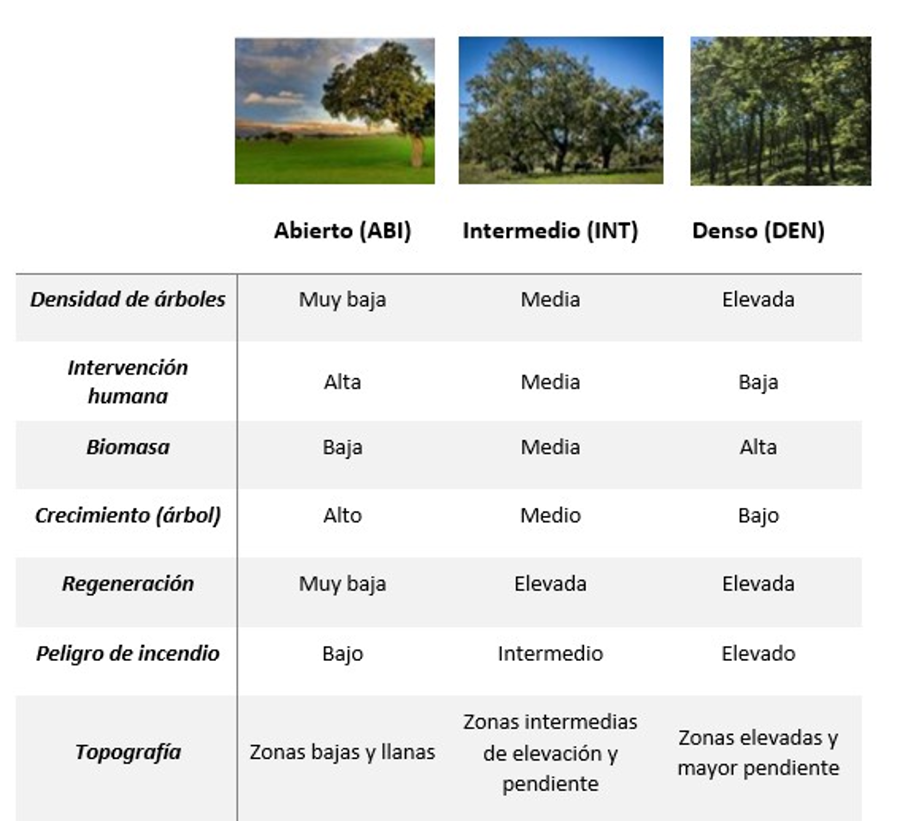
Figura 1.Características asociadas a encinares ibéricos dependiendo de su densidad de arbolado: Abierto (ABI); Intermedio (INT) y Denso (DEN).
Figure 1. Associated properties to Iberian holm oaks
forests based on tree density. Open (ABI); Medium (INT) and Dense
(DEN).
Datos y variables seleccionadas
Inventario Forestal
Nacional
El Inventario Forestal Nacional (IFN) establece parcelas
permanentes en una cuadrícula de 1 km2 en todo el territorio
forestal de España (Alberdi
et al. 2016). Las parcelas suelen
muestrearse siguiendo subparcelas concéntricas de 5, 10, 15 y 25 m de radio, en
las cuales se miden los diámetros y alturas de todos los árboles que presenten
un diámetro a la altura del pecho (DAP) superior a 7.5, 12.5, 22.5 y 42.5 cm,
respectivamente en cada subparcela (Durante et al. 2019).
Para el presente estudio se usó información del conjunto de datos del IFN2,
comprendido entre 1995 y 1997, y del IFN3 comprendido entre 2006 y 2008. Se
seleccionaron parcelas de clase “A”, y las subclases “1”, “3C” y “N”, las
cuales corresponden con parcelas analizadas durante el IFN2 y el IFN3, siendo
además comparables entre ellas. Esto nos permitió tener en cuenta las parcelas
que existen en ambas ediciones del inventario para calcular la productividad
forestal. Además, se seleccionaron las parcelas dentro del área de estudio
(Andalucía) clasificadas por tipo de bosque (Fig. 2).
En nuestro caso, la clasificación de tipos de bosque se realizó según la
densidad de arbolado de la parcela. Definiéndose como bosque abierto (ABI)
aquellas parcelas que presentan entre >20 y <50 árboles/ha, bosque
intermedio (INT) entre >150 y <350 árboles/ha, y bosque denso (DEN) con
un número de árboles igual o superior a 400 árboles/ha (Fig.
A1 del Anexo). Estos intervalos de densidad se hicieron en base al
número de pies en formaciones de bosque abierto y bosque denso en Q. ilex,
incluyéndose una clase intermedia que no solapase con los extremos. El IFN
clasifica morfo-estructuralmente las parcelas según sean “Bosques”, “Bosques de
plantación”, o “Bosques adehesados”. El 70% de las parcelas clasificadas como
“bosque adehesado” en el IFN corresponden con nuestra clasificación de “bosque
abierto”, el 81% de las parcelas clasificadas como “bosques” en el IFN
corresponden con nuestra clasificación de “bosque intermedio” o “bosque denso”,
mientras que los “bosques de plantación” son muy escasos (sólo un 2%). En
total, 1180 parcelas (563 ABI, 361 INT y 256 DEN) de encina coincidentes en
ambos IFNs fueron seleccionadas para su estudio en toda la región de Andalucía
(España).
Se usó el DAP para calcular la biomasa por árbol (kg·árbol-1)
mediante una ecuación alométrica descrita en Ruiz-Peinado
et al. (2012). La biomasa forestal (Mg·ha-1)
fue calculada para cada parcela como la suma de la biomasa individual de cada
árbol dentro de cada subparcela (Ruiz-Benito et al. 2014; Herraiz et al. 2023).
Se seleccionaron aquellas parcelas donde la especie de estudio (Q. ilex subsp.
ballota) es la especie dominante, es decir, presentaba más del 85% de la
biomasa de la parcela.
A partir de la biomasa del IFN3 y del IFN2 se calculó la
productividad como la diferencia de ambos valores dividido entre el tiempo
transcurrido entre ambos inventarios, estimándose como el crecimiento corriente
basado en los dos IFNs. También se calculó la densidad de árboles de cada
subparcela como la suma de árboles por ha.
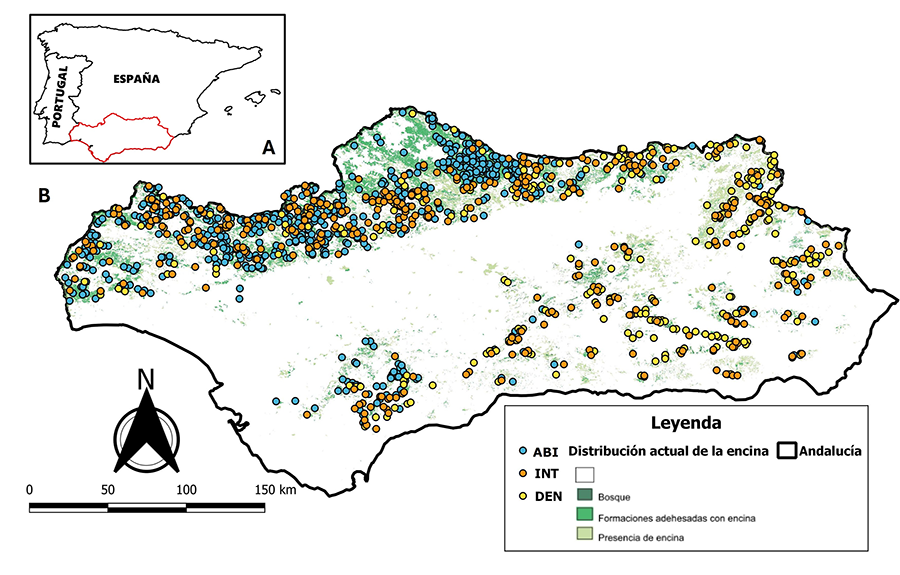
Figura 2. Área de estudio (Andalucía)
dentro de la península Ibérica (A); y ubicación de parcelas del tercer
Inventario Forestal Nacional (IFN3) clasificadas en Abierto (ABI), Intermedio
(INT) y Denso (DEN) dentro de Andalucía (B).
Figure 2. Study area (Andalucía) inside the península Ibérica (A); and location of third National
Forest Inventory (IFN3) plots classified as Open (ABI), Medium (INT) and Dense
(DEN) inside Andalucía (B).
Clima y topografía
También se tomaron datos de variables abióticas (climáticas,
topográficas, etc.) que podrían influir en los encinares. A partir de modelos
digitales de elevación en formato ráster con resolución de 30 m, y usando el
paquete “elevatr” en R, se calculó la pendiente (%) para todas las parcelas
como el máximo cambio de elevación entre una celda y sus celdas colindantes.
Por otra parte, se obtuvieron los datos de temperatura media anual (MAT) y de
precipitación (MAP) de la base de datos de WorldClim 1.2 (Hijmans 2012) para el mismo año de la toma de datos del IFN3
(2007). Además, se obtuvieron los datos de precipitación del mes más seco (DMP)
y temperatura del mes más seco (DMT), para calcular el índice de aridez
modificado de Martonne (AIm) atendiendo a la siguiente fórmula
(temperatura siempre en grados Celsius y precipitación en milímetros) (Stadler 2005):
AIm = {[MAP] / [MAT + 10]} + {[12 ×DMP] /
[DMT+10] / 2}
Dado que los valores más altos del AIm indican
una alta disponibilidad de agua, transformamos esta variable de la siguiente
manera: AI = 100-AIm.
La transformación de AI (100 − AIm) es completamente lineal y
no afecta a los resultados (Salazar-Zarzosa et
al. 2021). Así, los valores más altos del índice se corresponden con zonas
con menor humedad y viceversa.
También se obtuvieron los valores de precipitación media
anuales en Andalucía a partir de la Red de Información Ambiental de Andalucía (REDIAM),
para el periodo de tiempo del presente trabajo (2001-2023). A diferencia de
WorldClim, la REDIAM contiene todos los datos de precipitación para este
periodo con una mayor resolución temporal. Seguidamente, se realizó una
extracción de los valores de precipitación para cada parcela del IFN.
Índices de vegetación (IVs)
La obtención de los IVs se realizó mediante la plataforma
computacional basada en la nube Google Earth Engine (GEE) (Gorelick et al. 2017). Se usó una serie temporal de
datos de imágenes satelitales de Landsat 7 (Level 2, Collection 2, Tier 1), ya
corregidas atmosféricamente, que cubrían el periodo de estudio (2001-2023).
Este satélite fue seleccionado debido a su resolución temporal (15 días) y
espacial (30 m de tamaño de píxel). Los IVs seleccionados para el presente
trabajo fueron el índice de vegetación de diferencia normalizada (normalized
difference vegetation index, NDVI) y el índice de vegetación mejorado (enhanced
vegetation idex, EVI). El cálculo de los IVs se hizo siguiendo las fórmulas
siguientes, creando una composición con todas las imágenes para cada año, y
extrayendo el valor del píxel donde se ubica el centro de cada parcela del IFN.
NDVI = (NIR-R)/ (NIR+R)
EVI =G * ((NIR-R)/(NIR+C1*R-C2*B+L))
donde B, R y NIR corresponden a la reflexión espectral
propias del azul, rojo, e infrarrojo cercano, respectivamente. G es un factor
de ganancia con valor de 2,5. L es el ajuste de fondo de la cubierta vegetal
equivalente a 1. C1 y C2 son coeficientes de la resistencia de aerosol, con
valor de 6 y 7,5 respectivamente (Rouse et al. 1973;
Huete et al. 2002).
En un trabajo preliminar (Herraiz et
al. 2023) se comprobó que el valor del NDVI del píxel donde estaba
localizada la parcela se correlacionó fuertemente (R² = 0.93, P<0.001) con
el valor de NDVI de los 9 píxeles alrededor del punto de la parcela.
Seguidamente, se calculó la media anual de todos los años
del periodo de estudio, para todas las imágenes de la colección. Para estimar
cuantitativamente la salud vegetal a nivel de píxel y su dinámica en una serie
temporal, donde la sequía puede afectar, se empleó, además del NDVI y del EVI,
el índice de salud de la vegetación (vegetation health index, VHI). Se
incorporaron estos índices para cubrir las limitaciones ya conocidas del NDVI
en cuanto a la interferencia por la señal del suelo y en elevadas coberturas
vegetales (Huang et al.
2021; Mutanga et
al. 2023). Por su parte, el VHI fue
calculado para cuantificar e identificar la sequía sobre todo en plantaciones
agrícolas en base a la salud de la vegetación (Kogan 1995; Dalezios et al. 2014).
Es un índice calculado a partir del NDVI y de la temperatura superficial
terrestre. Todos ellos (NDVI, EVI y VHI) tienen un rango de valores entre -1 y
1, representando un mejor estado de la vegetación cuanto mayor sea el valor (Kogan 1995). El VHI mide el estado de la salud de la
vegetación partiendo del propio NDVI, e incluyendo en su fórmula la temperatura
superficial de la tierra (LST) para tener en cuenta las condiciones térmicas.
Para obtener el VHI, primero se calcula el índice de condición de la vegetación
(VCI) y después el índice de condición térmica (TCI) de la siguiente manera:
VCI = (NDVI - NDVImin)/(NDVImax
-NDVImin)
TCI = (LSTmax
- LST) / (LSTmax - LSTmin); siendo LST el valor de
temperatura de la superficie, LSTmax el valor máximo del mismo y LSTmin
el mínimo.
VHI
=a*VCI + (1- a)*TCI,
siendo “a” el coeficiente de
la contribución de cada índice (VCI y TCI) entre 0 y 1. En este caso, se
consideró el valor de 0.5 para ambos.
Para el NDVI se obtuvieron los valores medios (NDVImedio),
mínimos (NDVImin) y máximos (NDVImáx) anuales, necesarios
en el cálculo del VHI, y porque representan las dinámicas fenológicas a lo
largo del año. De igual forma para el EVI se calcularon los valores medios (EVImedio),
mínimos (EVImin) y máximos (EVImáx). En ambos IVs, se
consideran estas tres métricas por su importancia a la hora de monitorizar la
vegetación y por su capacidad de diferir entre tipos de bosque (Torres et al. 2021).
En resumen, se obtuvo una base de datos con todas las
variables medidas en campo a partir del IFN, y los IVs correspondientes al
mismo año de muestreo (2007).
Para el tercer objetivo, se obtuvo una base de datos
multitemporal con los valores de todos los IVs calculados anualmente (valores
medios, mínimos y máximos anuales) y la precipitación anual acumulada para cada
parcela en el periodo de tiempo 2001-2023.
Análisis de datos
Para evaluar las
diferencias entre los distintos tipos de bosque (ABI, INT, y DEN) (Objetivo
1), con el paquete “stats” de R base, se realizó una prueba no paramétrica
de Kruskal-Wallis (K-W) de las variables de interés (aridez, precipitación,
elevación, Biomasa IFN3, Productividad, Nº de especies matorral IFN3 y NDVImin),
dado que los datos no seguían una distribución normal. Se usó el valor mínimo
de uno de los IVs para tratar de disminuir la influencia de pastos o matorrales
en la señal espectral de las parcelas, sobre todo en las de bosque abierto (Wang y Tenhunen 2004; Hoek van Dijke et al. 2023). Posteriormente, se aplicó una prueba de
comparación de medias por pares (test de Wilcoxon) para saber qué grupos eran
significativamente diferentes entre sí. Además, con la finalidad de saber qué
variables son diferenciadoras del tipo de bosque, se realizó un modelo
multinomial con los tipos de bosque como variable respuesta y las variables
mencionadas anteriormente como predictoras. Para ver si las variables de
nuestra base de datos (IFN e IVs) estaban correlacionadas, se realizó un
análisis de correlación (test de Spearman) con el paquete “corrplot” (Fig. A2 del
Anexo), y posteriormente un análisis de
componentes principales (PCA) usando el paquete “FactoMineR” (Figs. A3 – A5
del Anexo). Las variables incluidas en el
PCA fueron: aridez, precipitación, pendiente, biomasa IFN3, productividad, EVImin,
NDVImin, VHI. Los mínimos de los IVs fueron seleccionados por el
mismo motivo que anteriormente (eliminar ruido del resto de vegetación). Se
hizo además un test de ANOVA (paquete “stats”) entre los datos de todas las
variables de los tres tipos de bosque para buscar diferencias significativas
entre sus medias en el primer y segundo componente.
Posteriormente, para la
consecución del Objetivo 2 se realizaron modelos lineales generalizados
(GLMs) (función glm, paquete “stats”) donde la variable dependiente fueron los
IVs (NDVImedio, NDVImax, NDVImin, EVImedio,
EVImax, EVImin y VHI), y las variables independientes
fueron: precipitación, aridez, biomasa del IFN3 y pendiente. Los GLMs se
llevaron a cabo marcando la familia “gamma” por el tipo de distribución que
seguían los datos de los IVs. Para ajustar la presencia de valores negativos se
escalaron todos los valores de los IVs mínimos, para poder usar así dicha
distribución en todos los GLMs. Las variables independientes fueron
seleccionadas evitando la correlación entre ellas tras el correspondiente
análisis (Fig.
A2 del Anexo) y seleccionando
aquellas que intervienen de manera más directa en la estructura y dinámica de
los bosques (Bolstad et al. 2018). Los GLMs son métodos estadísticos flexibles para analizar las
relaciones ecológicas, ya que tienen en cuenta el comportamiento interactivo de
variables que pueden estar vagamente representadas por distribuciones
gaussianas (lineales) clásicas. Para conocer el rendimiento de los modelos se
usaron diferentes métricas e indicadores como el criterio de información de
Akaike corregido (AICc), el R2 ajustado (R2 adj.) o el
pseudo-R2 de Nagelkerke (Tablas A2-A4 del Anexo). Además, se realizó una prueba de verosimilitud
para comparar nuestros modelos y determinar si nuestras variables tienen mayor
poder explicativo que un modelo vacío (IVs ~ 1). A todos los modelos se les
realizó un ANOVA, usando la prueba de Chi cuadrado, para compararlos y conocer
así si nuestros modelos eran significativos. Los valores de AICc de estos
modelos vacíos también fueron comparados con los de los modelos reales para
detectar los modelos con mejor rendimiento según esta métrica de rendimiento
(menor valor de AICc).
Para abordar el Objetivo 3, relacionado con la
variabilidad interanual de los IVs por tipo de bosque, se seleccionaron los
índices con mejor rendimiento en los GLMs, es decir, aquellos cuyas métricas de
rendimiento son mejores (menor AICc en comparación con su modelo vacío y mayor
R2 adj. y pseudo-R2 de Nagelkerke). A partir de los
promedios anuales de los IVs seleccionados (NDVImax y EVImax),
se midió la correlación entre estos índices con el coeficiente de correlación
de Spearman (ρ). Posteriormente, calculamos las tendencias (pendiente) de estos
IVs para una serie temporal de 23 años (2001-2023) mediante el test de
Mann-Kendall (Mann 1945; Kendall 1975) para cada tipo de parcela del IFN (paquete
“Kendall”). Una vez calculadas las tendencias, fueron representadas
espacialmente (paquetes “sf” y “ggplot2”) (Fig. 5).
Para conocer la respuesta en el índice de verdor de cada tipo de bosque frente
a periodos de baja precipitación o sequía, se realizaron análisis de
correlación de Pearson entre los IVs y los valores de precipitación anual.
Además, se realizó una prueba de correlación cruzada que consiste en analizar
si la correlación entre las variables se da con un efecto de retardo, temprano
o tardío (Vio y
Wamsteker 2001); en este caso, analizamos
si el retardo del valor de los IVs ocurría hasta 2 años antes o después de cada
valor de precipitación.
Todos los análisis fueron llevados a cabo en el entorno de
programación de R (versión 4.2.2).
Resultados
Diferencias entre los tipos de bosque de encina
Se observaron diferencias significativas (p < 0.05) entre
los tipos de bosque para todas las variables estudiadas (Fig.
3), excepto para la precipitación (K-W p-value = 0.06).
Concretamente, las mayores diferencias fueron para la aridez, la elevación, la
pendiente, la productividad, el número de especies del matorral y el NDVImin.
Los bosques DEN son los que mostraron una menor aridez (Fig.
3A), mayor precipitación, elevación y pendiente (Figs.
3B, C y D), mayor biomasa y productividad (Figs.
3E y F), mayor número de especies de matorral (Fig.
3G), y mayor NDVImin (Fig. 3H),
mientras que para el ABI ocurrió lo opuesto. Los valores de los bosques INT se
encuentran entre los DEN y ABI. También hubo variables que muestran diferencias
significativas sólo entre dos tipos de bosque, y no entre los tres. Es el caso
de la biomasa (Fig. 3E), donde los valores más
altos se dieron para el DEN diferenciándose significativamente del INT y del
ABI, respectivamente.
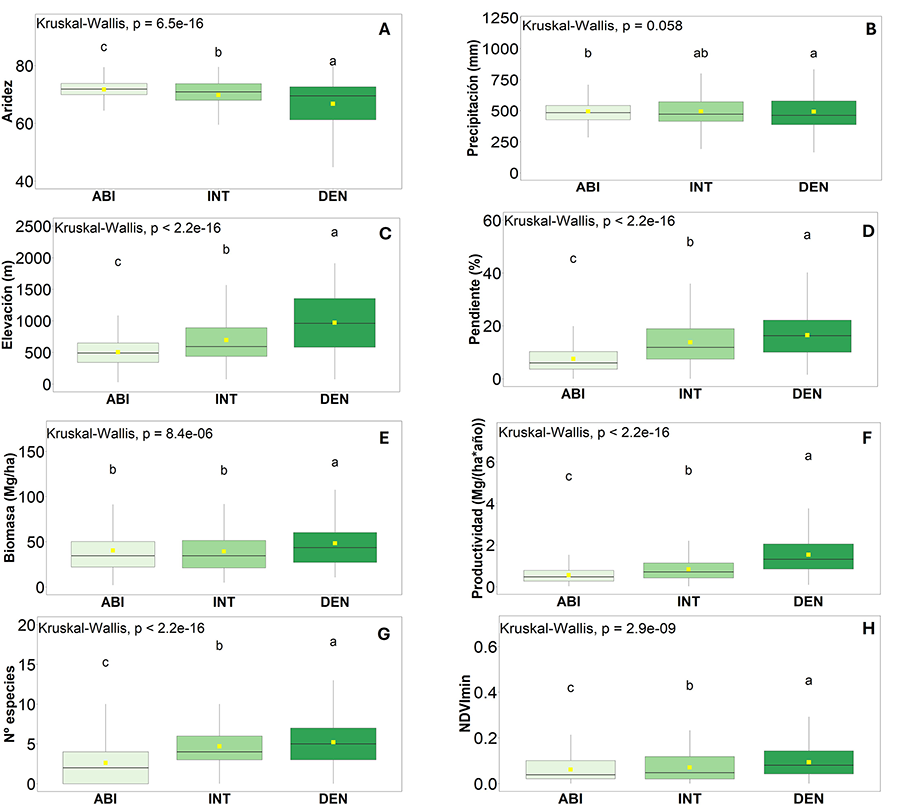
Figura 3. Comparación
de las variables de estudio, bióticas y abióticas, entre los diferentes tipos
de bosque: Abierto (ABI), Intermedio (INT) y Denso (DEN). Límite inferior de la
caja como primer cuartil (Q1), límite superior como tercer cuartil (Q3), línea
divisoria como mediana o segundo cuartil (Q2) y punto amarillo como la media.
Las barras indican la desviación estándar. Diferentes letras indican una
diferencia estadística (P<0.05) entre grupos.
Figure
3. Comparison of biotic and abiotic study variables
between forest types: Open (ABI), Intermedio (INT) y Denso (DEN). Lower
boundary of the box as first quartile (Q1), upper boundary as third quartile
(Q3), dividing line as median or second quartile (Q2) and yellow dot as the
mean. The whiskers indicate the standard deviation. Different letters indicate
a statistical difference (P<0.05) between groups.
En el análisis de ordenación (PCA), la varianza
absorbida por el componente 1 y 2 fue de un 30.4% y un 17.1%, respectivamente (Fig. A3 del Anexo). Las variables más relacionadas
con el componente 1 fueron la elevación y la aridez (Fig.
A4 del Anexo), mientras que para el componente 2 fueron los mínimos del
NDVI y EVI (Fig. A4 del Anexo). Se observó que
no existen diferencias significativas en las medias de los tres grupos, tampoco
en el componente principal 1 ni en el componente principal 2, aunque se puede
ver que el espacio ocupado es mayor en los bosques DEN frente al ABI e INT, mostrando
los primeros una mayor variabilidad.
Modelos lineales generalizados
Los resultados de los GLMs realizados por tipo de bosque
mostraron que, de las tres métricas de cada índice de verdor además del VHI, la
mejor relación se da en el NDVImax y EVImax (Tabla 1, Tablas A2-A4 del
Anexo), siendo los modelos significativos para los tres tipos de bosque (p-value
< 0.05). Independientemente del tipo de bosque, esta métrica es la que mejor
se relacionó con las variables biofísicas (R2 adj. NDVImax
medio = 0.17; R2 adj. EVImax medio = 0.15). Tras los
valores máximos, es la media la que presentó un mejor rendimiento en los
modelos.
Con respecto a los coeficientes significativos de los
modelos (Tabla 1, Tabla
A1 del Anexo), el “intercept” fue significativo en todos los
modelos de todos los tipos de bosque, excepto en los valores mínimos. Las
únicas variables que aparecieron como significativas en algunos modelos fueron
la biomasa (7 de 21) y la aridez (2 de 21) (Tabla A1
del Anexo). El resto de variables (precipitación y pendiente) no presentaron
significancia en ninguno de los modelos.
Tabla 1.
Aportación de las variables a los GLMs con mejor rendimiento para los tres
tipos de bosque (ABI, abierto; INT, intermedio; DEN, denso). Grado de
significancia representado por *** (p-value <0.001), ** (0.001 < p-value
< 0.01), ** (0.001 < p-value < 0.01), * (0.01 < p-value
< 0.05). NDVImáx: Índice de Diferencia Normalizada de la
Vegetación (máximo anual); EVImáx: Índice de Vegetación Mejorado
(máximo anual).
Table
1. Contribution of
the variables to the GLMs with best performance for the three types of forests
(ABI, open; INT, intermediate; DEN, dense). Degree of significance represented
by: *** (p-value < 0.001), ** (0.001 < p-value < 0.01),
* (0.01 < p-value < 0.05). NDVImax: Normalized
Difference Vegetation Index (annual maximum); EVImax: Enhanced
Vegetation Index (annual maximum).
|
|
Tipo bosque
|
Biomasa
|
Precipitación
|
Pendiente
|
Aridez
|
R2 adj.
|
|
NDVImáx
|
ABI
|
0.0005
|
-2.65E+09
|
-0.0019
|
-0.0013*
|
0.108
|
|
INT
|
0.0006
|
-3.50E+09
|
-0.0002
|
-0.0031
|
0.200
|
|
DEN
|
0.0006
|
1.88E+09
|
0.0004
|
-0.0026
|
0.185
|
|
EVImáx
|
ABI
|
0.0008
|
-4.46E+09
|
-0.0035
|
-0.0017
|
0.112
|
|
INT
|
0.0009
|
-4.92E+09
|
-0.0002
|
-0.0049
|
0.180
|
|
DEN
|
0.0009
|
4.11E+09
|
0.0005
|
-0.0040
|
0.171
|
Análisis multitemporal
de IVs
Se realizó un análisis multitemporal para el periodo
2001-2023 de sus valores anuales por cada tipo de bosque, superponiendo a la
vez los valores anuales de precipitación media (Fig. 4).
El NDVImax mostró unos valores mayores en ABI durante toda la serie
con respecto a los demás tipos de bosque, pero siempre manteniendo las mismas
tendencias temporales entre los tres tipos de bosque (Fig.
4). El NDVImax mantuvo valores inferiores a los del EVImax
en todo el periodo de tiempo. Por su parte, el EVImax mostró más
variación entre los valores de los distintos tipos de bosque a lo largo de la
serie temporal, aunque para ambos índices, los valores mayores correspondieron
con el bosque ABI. El NDVImax presentó un coeficiente de correlación
de Pearson no significativo con la precipitación de 0.22 (p-value =
0.07), mientras que el EVImax presentó un coeficiente de correlación
de Pearson significativo y mayor (0.28; p-value = 0.02). También se
realizó un análisis de correlación cruzada para analizar si la precipitación
tenía un efecto retardado en los IVs. Tanto el NDVImax como el EVImax
presentaron una mayor correlación con el retardo = 0 (Fig.
A6 del Anexo).
En relación con los valores de precipitación, hay años donde
se dieron eventos de sequía extremos, como en los años 2015, 2017 o 2023,
destacando el año 2005. Para los años de sequía, los valores de precipitación
de los tres tipos de bosque son prácticamente similares.
Tomando NDVImax y EVImax como
indicadores de referencia según los resultados de los GLMs, la tendencia de los
índices en el periodo 2001-2023 para las parcelas según el tipo de bosque fue
representada espacialmente (Fig. 5). El EVImax
presenta una tendencia negativa casi en la totalidad de las parcelas de
los tres tipos de bosque. El NDVImax muestra zonas donde se
concentra el signo de la tendencia (ya sea positiva o negativa). Los puntos más
al este presentan una tendencia positiva sobre todo en el DEN y en el INT,
mientras que los del norte de Andalucía en el tipo ABI muestran todos una tendencia
negativa.
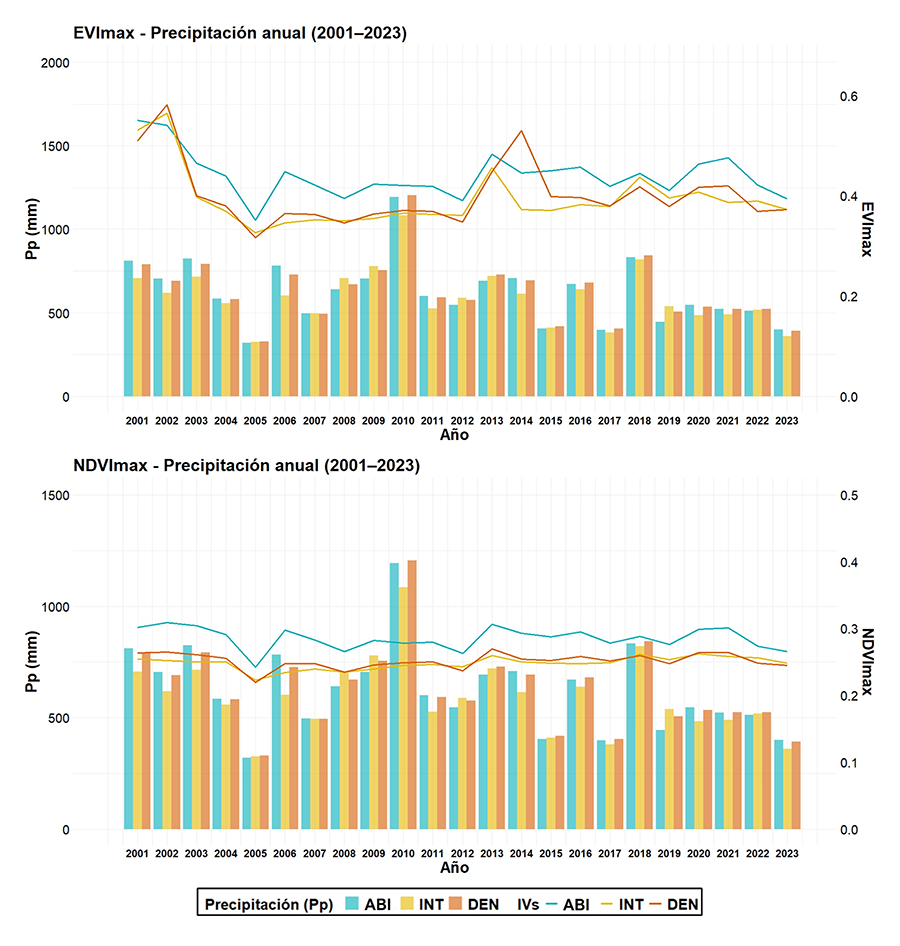
Figura 4. Análisis multitemporal de NDVImax,
EVImax y de la precipitación media por tipo de bosque clasificado en
Abierto (ABI), Intermedio (INT) y Denso (DEN) entre 2001 y 2023.
Figure 4. Multitemporal analysis of NDVImax, EVImax and
mean precipitation by forest type classified as Open (ABI), Medium (INT) and
Dense (DEN) between 2001 and 2023.
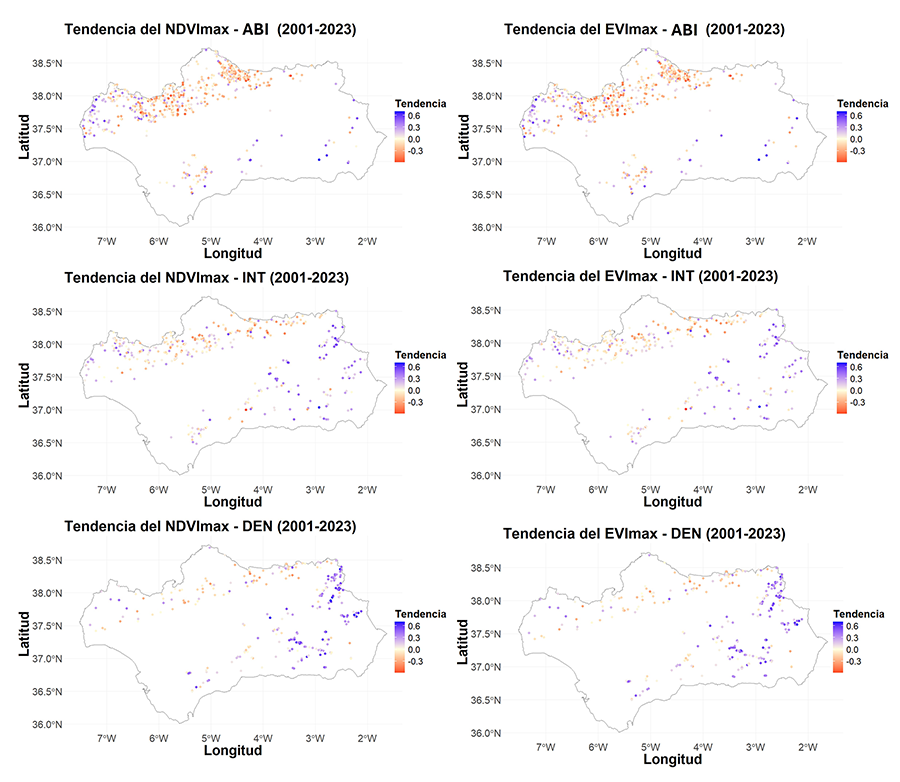
Figura 5. Representación espacial de la
tendencia temporal del NDVImax y del EVImax por parcela
del IFN de cada tipo de bosque clasificado en Abierto (ABI), Intermedio (INT) y
Denso (DEN) entre 2001 y 2023.
Figure
5. Spatial representation of the temporal trend of
the NDVImax and EVImax by forest type classified as Open
(ABI), Medium (INT) and Dense (DEN) between 2001 and 2023.
Discusión
Diferencias entre los tipos de bosque según su
densidad
Nuestros resultados muestran que los bosques mediterráneos
de encina en Andalucía presentan diferencias en su estructura y dinámica
funcional. Aunque la densidad de arbolado no es la única indicadora de la
gestión, es cierto que los bosques abiertos suelen haber sufrido un aclarado
mayor a lo largo de la historia y por lo tanto una mayor intervención humana (Pulido et al. 2010). Los bosques densos tienen una
biomasa superior que los bosques intermedios, y todavía mayor que los abiertos,
cómo es lógico debido a su mayor densidad de árboles, mostrando además una
mayor productividad (Fig. 3). Los bosques más
densos suelen ubicarse en áreas de mayor pendiente y altitud, donde la
implantación de sistemas agrícolas ha resultado históricamente más difícil.
Estas zonas, por su inaccesibilidad o menor aptitud para la agricultura
intensiva, han sido menos transformadas por la actividad humana, lo que ha
favorecido la conservación de coberturas forestales más cerradas. Aunque podría
pensarse que estas áreas son menos productivas —lo cual justificaría su menor
uso agrícola—, diversos estudios han mostrado que en ciertos contextos
presentan niveles de productividad relativamente altos (Laamrani
et al. 2014; Spasojevic et al. 2015). No
obstante, la relación entre productividad y topografía tiende a ser indirecta (Helman et al. 2017), ya que factores como la retención
de agua, la exposición solar o la acumulación de materia orgánica —todos ellos
influenciados por la topografía— son los que afectan realmente la
productividad, más que la altitud o pendiente por sí solas. La edad de la
encina, al igual que la densidad del arbolado, es un factor determinante en la
productividad (Wang et al. 2011). Esto se debe a que,
en zonas de menor pendiente, donde se suelen encontrar los bosques más abiertos
y/o dehesas, los árboles tienden a ser más longevos en comparación con áreas
montañosas de pendiente más pronunciada, donde además los árboles se cortan
para la producción de leña o carbón (Gea-Izquierdo
et al. 2008; Salazar-Zarzosa et al. 2021).
La aridez es una variable relevante para diferenciar entre
los tres tipos de bosque, presentando valores más altos en los encinares
abiertos (ABI) y disminuyendo gradualmente a medida que aumenta la densidad del
arbolado. Esta distribución tiene sentido si consideramos que los ABI se
localizan predominantemente en zonas más bajas, caracterizadas por menores
precipitaciones y temperaturas más elevadas, es decir, en contextos
climáticamente más áridos. No es que la estructura abierta de estos bosques
cause una mayor aridez, sino que estos tipos de bosque se desarrollan en
entornos más secos. Además, la gestión más intensiva en estos sistemas puede
reducir la cobertura del sotobosque, lo que limita aún más la capacidad de
retención de humedad del ecosistema (Díaz et al. 2021). Aunque
en algunos contextos una menor densidad de arbolado puede reducir la
competencia por recursos como el agua —lo cual resulta beneficioso en entornos
áridos—, también se ha observado que una mayor separación entre individuos
puede incrementar la vulnerabilidad de los ecosistemas forestales frente al
cambio climático, especialmente en lo que respecta a la pérdida de conectividad
estructural y funcional, la mayor exposición de los suelos y la reducción del
microclima generado por la cubierta arbórea (Young et al.
2023). Esta vulnerabilidad no se limita exclusivamente a los bosques
abiertos, pero en ellos puede ser más pronunciada debido a la menor resiliencia
estructural frente a perturbaciones externas. Este patrón también se observa a
través del EVImin, aumentando el índice de verdor a medida que
aumenta la densidad del arbolado. Además, los valores mínimos permitieron
distinguir significativamente los diferentes tipos de bosque según su señal
espectral como demuestran nuestros resultados (Fig. 3H).
Al tratarse de valores mínimos, podemos disminuir la señal de especies no
arbóreas que pueden tener picos elevados de verdor e influir en los datos (Wang y Tenhunen 2004). Así lo vemos en los resultados obtenidos (Fig. A7 del Anexo). En este contexto, y con el
objetivo de mitigar los efectos del cambio global, concretamente con anomalías
climáticas provocadas por el cambio climático, existen alternativas para
reducir o mitigar la pérdida de biodiversidad y de los servicios ecosistémicos
asociados a este tipo de ecosistemas. Entre otras, conservar un sotobosque que
minimice la pérdida de agua (Rasoanaivo et al. 2024)
o crear zonas abiertas alternadas con áreas de alta densidad de matorral, que
se vayan rotando de manera periódica (Oksuz et al. 2020).
El uso de índices espectrales de vegetación (IVs)
para caracterizar los tipos de bosque
En general, las diferencias
entre los tres tipos de bosque fueron explicadas por la relación entre los IVs
y las variables tanto abióticas como bióticas medidas en las parcelas del IFN.
Según los modelos lineales generalizados, la variabilidad de los valores
máximos de NDVI y EVI se explica parcialmente por la aridez y la biomasa en más
de un 15% (R2 adjust. > 0.15) excepto para el bosque abierto
donde sólo explican un 10 % de estos valores (R2 adjust. > 0.10). La
significancia muestra que tan sólo la aridez es significativa en un tercio de
los modelos. Estos resultados coinciden además con el análisis de componentes
principales en el que se utilizan estas mismas variables, donde las medias de
los tipos de bosque no son significativamente diferentes cuando se incluyen
estas variables en dicho análisis (Fig. A5 del
Anexo). Por lo tanto, en base a esto y a las métricas de rendimiento de los
GLMs (Tablas A2-A4 del Anexo), de todas las
métricas analizadas (media, máximos y mínimos) se pueden considerar el NDVImax
y EVImax, como los indicadores más idóneos a la hora de describir
ciertas características biofísicas de los tipos de bosque estudiados (Freitas et al. 2005).
Nuestros resultados muestran
que, a pesar de que la especie dominante es la encina, la respuesta espectral
basada en los IVs de los tres tipos de bosque dependería en parte, a falta de
análisis más específicos, de la densidad de árboles y de determinadas variables
ambientales de cada parcela estudiada del IFN (Herraiz et al. 2023).
Estos efectos podrían reflejarse en los valores más altos de ciertos índices de
verdor (valores máximos del EVI y NDVI; Fig. A7 del
Anexo) observados en los bosques ABI en comparación con los DEN. Sin embargo,
es importante matizar que no todos los índices siguen el mismo patrón; por
ejemplo, en el caso del EVImin, ya se ha señalado previamente que
presenta una tendencia distinta, lo que subraya la necesidad de considerar cada
métrica por separado al interpretar estas diferencias, ya que podrían estar
condicionadas por otros factores además de la densidad de arbolado. Estos
efectos se tendrán en cuenta en futuros trabajos mediante la aplicación de
filtros correctores.
Esta respuesta diferente tiene, por tanto, un efecto en la
provisión de diferentes funciones y servicios ecosistémicos de este tipo de
bosques, como reservas de carbono, productividad o floración, así como en la
biodiversidad que albergan (Bölöni et al. 2021; Bastias et al. 2025).
Por lo tanto, aunque los datos obtenidos por satélite no pueden reemplazar a
los datos de campo, sí los complementan, lo que aporta una serie de importantes
ventajas. Nuestros resultados confirman, además, el uso de datos de
teledetección como método alternativo para la realización de mapas de cobertura
del suelo con más detalle (Wang
et al. 2023).
El análisis de las tendencias de índices
espectrales de vegetación como herramienta de monitoreo del estrés climático en
los tipos de bosque
Nuestros resultados se alinean en gran medida con aquellos
que usan IVs obtenidos a partir de datos de satélite para analizar la evolución
de las masas vegetales con el tiempo. Dichos estudios han puesto de manifiesto
un predominio de las tendencias positivas en el reverdecimiento tanto a escala
de Andalucía (Gutiérrez
Hernández 2022; Herraiz et al. 2025), como de la península ibérica (Vicente-Serrano et
al. 2020), continental (Cortés et al. 2021) y global (Chen et al. 2019). Sin
embargo, la mayoría de estos estudios se basan en el análisis de las tendencias
de IVs, principalmente NDVI, a nivel de píxel, con una resolución espacial
(tamaño de píxel) baja (MODIS a 250m y NOAA-AVHR a 1km). En nuestro caso,
proporcionamos un análisis espacial y temporal con más detalle del
reverdecimiento basado en la gestión de las parcelas del IFN (~30m), lo cual
ofrece grandes ventajas al combinar la precisión y detalle del tamaño de la
parcela con la información espectral de la estructura y composición del tipo de
bosque a lo largo del tiempo.
Los resultados del análisis multitemporal para un periodo de
23 años (2001-2023) muestran que existe una relación entre la dinámica de los
IVs y la precipitación para cada tipo de bosque (Aksoy y Sertel 2021).
Esto demuestra que la precipitación juega un papel importante como reguladora
de los IVs, pudiendo ayudar a predecir o explicar la respuesta a futuros
eventos de cambio climático (Li
et al. 2018). De manera específica, existe
una mayor correlación entre la precipitación y el EVI, lo cual puede deberse al
parámetro corrector de la señal del suelo que introduce el cálculo de este
índice. Aun así, es necesario un avance más profundo en la aplicación de
filtros y correcciones que permitan eliminar la reflectancia del suelo, sobre
todo en zonas más abiertas. Además, el hecho de que en los años de mayor sequía
(por ejemplo 2005) se produzca un descenso notable de los valores de los IVs
para los tres tipos de bosque (Fig. 4), y sobre
todo para el EVI (años 2015 y 2017, Fig. 4), con
una recuperación tras el periodo de sequía (2018, Fig. 4),
indica que el análisis de series temporales de IVs permite una alerta temprana
de las respuestas de la vegetación a los impactos de eventos climáticos
extremos, como la sequía (Zellou et al. 2023; Kladny et al. 2024). Para abordar más en profundidad estas cuestiones
en futuros trabajos, se podrían seleccionar aquellas parcelas cuya biomasa no
varía sustancialmente de forma natural y asociar de manera más directa un
cambio en éstas con el estrés climático. Respecto a los valores más altos del
bosque abierto, pueden venir condicionados por la existencia de otras cubiertas
de vegetación como pasto o matorral disperso, que podría influir en la señal
espectral de los IVs, lo cual requiere de un análisis más detallado (por ejemplo,
usando los valores mínimos de los IVs).
Según nuestros resultados existen “hotspots” de parcelas con
tendencia positiva y “coldspots” de parcelas con tendencia negativa en base al
reverdecimiento dependiendo del tipo de bosque. Pueden venir derivados de la
diferencia de manejo entre los tipos de bosque. Las parcelas de tipo ABI
presentan un mayor control que limita el incremento de masa forestal,
sucediendo todo lo contrario en los INT y DEN (Bartušková et al. 2015).
En estos últimos, el aumento de la masa forestal es producido, entre otros
factores, por el abandono, lo que hace que la masa forestal aumente y la
tendencia sea positiva (Herraiz et al. 2025).
Dependiendo del tiempo y/o grado de este abandono, podrían crearse zonas donde
la tendencia sea más acentuada.
Integración de la información de los inventarios
forestales con la teledetección satelital para mejorar el monitoreo de los
bosques mediterráneos
En general, nuestro estudio
demuestra que el uso combinado de parcelas del IFN y de IVs obtenidos a partir
de imágenes satelitales ofrece ventajas e inconvenientes en la evaluación y
monitoreo de los encinares en Andalucía. Entre las ventajas, el uso de parcelas
del IFN permite obtener datos precisos y detallados in-situ sobre la
estructura y composición del bosque en un momento específico del tiempo, lo que
facilita un análisis riguroso a nivel local. Sin embargo, los muestreos en las
parcelas de inventario son costosos y requieren mucho tiempo, lo que limita su
cobertura espacial y temporal. En contraposición, los IVs derivados de imágenes
satelitales se relacionan de manera aceptable con los datos de campo, siendo
extrapolables a una escala temporal mayor, en este caso mediante un análisis
interanual. Estos análisis multitemporales nos permiten además monitorear
grandes extensiones de terreno de manera rápida y frecuente proporcionando
información sobre la salud y dinámica de la vegetación a escala regional o
global. Por lo tanto, ambos enfoques se complementan, permitiendo una visión
más completa cuando se utilizan conjuntamente (Nesha et al. 2022).
No obstante, las imágenes
satelitales pueden presentar limitaciones en la precisión espacial o la
resolución temporal, además de no ofrecer detalles específicos sobre la
estructura interna del bosque, lo cual requiere del uso de imágenes con un
mayor detalle. En este sentido, es necesario explorar la componente temporal de
manera más fina, por ejemplo, a través de análisis intra-anuales, lo cual es
factible mediante el uso de sensores satelitales con una resolución temporal
más alta. Además de proporcionar información detallada, una mayor resolución
espacial permitiría estudiar la fenología o estacionalidad de los tipos de
bosque, detectando su capacidad de respuesta y recuperación ante eventos
extremos como la sequía.
Conclusiones
A la luz de los resultados obtenidos en este estudio,
podemos considerar válidas las tres hipótesis planteadas. Determinadas
variables biofísicas (estructurales, funcionales y de composición) medidas en
campo (a través de las parcelas del IFN) tienen un rol muy importante en la
estructura y composición de los tipos de bosque de encina. Además, la señal
espectral de los IVs está condicionada, principalmente, por la aridez. Por lo
tanto, aunque los datos satelitales no sustituyen a los datos de campo, sí complementan
el análisis, facilitando la evaluación de cómo factores como la densidad de
árboles y la gestión pueden influir en los servicios ecosistémicos de este tipo
de bosques. Nuestro estudio confirma el potencial de la combinación de los
datos de las parcelas del IFN a escala espacial fina, con los valores de los
IVs obtenidos a partir de series temporales de imágenes de satélite, para
informar sobre la dinámica forestal y los tipos de encina en Andalucía. Este
enfoque integrado proporciona un análisis más detallado de la dinámica
forestal, y establece una base sólida para futuros estudios orientados a
examinar con mayor precisión, espacial y temporal, la capacidad de recuperación
de los bosques mediterráneos ante eventos extremos.
Contribución de los autores
Velasco-Rodríguez, A.: Análisis formal,
conceptualización, curaduría de datos, metodología, software, validación,
redacción del borrador original, revisión y edición. Villar, R.:
Administración del proyecto, adquisición de fondos, conceptualización,
metodología, recursos, supervisión, validación, redacción, revisión y edición. Herraiz,
A.D.: Curaduría de datos, software, recursos, redacción, revisión y
edición. Salazar-Zarzosa, P.: Curaduría de datos, software, recursos,
redacción, revisión y edición. Castro, J.: Curaduría de datos,
metodología, redacción, revisión y edición. Arenas-Castro, S.:
Administración del proyecto, adquisición de fondos, conceptualización,
metodología, recursos, supervisión, validación, redacción, revisión y edición.
Disponibilidad de datos y código
Los datos originales de este trabajo se encuentran
depositados en el repositorio Zenodo.org y son accesibles a través del
siguiente enlace: https://doi.org/10.5281/zenodo.15100338.
Financiación, permisos requeridos, potenciales
conflictos de interés y agradecimientos
Este estudio está financiado por el proyecto FORMEDY
(TED2021-131722B-IOO), financiado por el MCIN/AEVl0.13039/501100011033 y por la
Unión Europea ''NextGenerationEU "/PRTR. Plan de Recuperación,
Transformación y Resiliencia - Unión Europea -NextGenerationEU. Así mismo, es
parte del proyecto FOR_FUN (PID2020-115809RB-I00) del MEC (Gobierno de España)
y fondos FEDER. Esta investigación también contó con el apoyo del proyecto
BOSQUES (BIOD22_0003_21_PPCB) a través del “Plan Complementario de I+D+i en el
área de Biodiversidad (PCBIO)” financiado por la Unión Europea en el marco del
Plan de Recuperación, Transformación y Resiliencia - NextGenerationEU y por la
Junta de Andalucía. Parte de este estudio fue llevado a cabo dentro del
contexto del Trabajo Fin de Máster realizado por Joselyn Castro en el Máster en
Geomática, Teledetección y Modelos espaciales aplicado a la gestión forestal
(Universidad de Córdoba, España).
Referencias
Aksoy, S., Sertel, E. 2021. Comparison of Landsat and Modis derived vegetation health indices
for drought monitoring using google earth engine platform. In: Proccedings
of the International Symposium on Applied Geoinformatics 2021, Riga, Latvia,
(ISAG2021). https://doi.org/10.15659/isag2021.12604
Alberdi, I., Sandoval, V., Condés,
S., Cañellas, I., Vallejo, R. 2016. El Inventario Forestal Nacional español,
una herramienta para el conocimiento, la gestión y la conservación de los
ecosistemas forestales arbolados. Ecosistemas 25: 88-97. https://doi.org/10.7818/ECOS.2016.25-3.10
Alcaraz-Segura, D., Baldi,
G., Durante, P., Garbulsky, M.F. 2008. Análisis de la dinámica temporal del
NDVI en áreas protegidas: tres casos de estudio a distintas escalas espaciales,
temporales y de gestión. Ecosistemas 17(3):108-117. Retrieved from https://www.revistaecosistemas.net/index.php/ecosistemas/article/view/82
Anderegg, W.R.L., Trugman, A.T., Badgley, G., Anderson, C.M., Bartuska, A.,
Ciais, P., Cullenward, D., et al. 2020. Climate-driven risks to the climate
mitigation potential of forests. Science 368. https://doi.org/10.1126/science.aaz7005
Araque Jiménez,
E., Sánchez Martínez, J.D. 2009. Repoblación forestal en Andalucía. Universidad
de Jaén (ed.). Jaén, España.
Bartušková, A., Doležal, J.,
Janeček, Š., Lanta, V., Klimešová, J. 2015. Changes in biomass
allocation in species rich meadow after abandonment: Ecological strategy or
allometry? Perspectives in Plant Ecology, Evolution and Systematics 17:
379-387. https://doi.org/10.1016/j.ppees.2015.06.003
Bastias, C.C., Rodríguez Castilla,
G., Salazar Zarzosa, P., Díaz Herraiz, A., González Herranz, N., Ruiz-Benito,
P., Barrón, V., et al. 2025. Differential aridity-induced
variations in ecosystem multifunctionality between Iberian Pinus and Quercus
Mediterranean forests. Ecological Indicators 173, 113411. https://doi.org/10.1016/j.ecolind.2025.113411
Bento, V.A., Gouveia, C.M., DaCamara, C.C., Trigo, I.F. 2018. A climatological assessment of drought impact on vegetation health
index. Agricultural and Forest Meteorology 259: 286-295. https://doi.org/10.1016/j.agrformet.2018.05.014
Bölöni, J., Aszalós, R., Frank, T., Ódor, P. 2021. Forest type matters:
Global review about the structure of oak dominated old-growth temperate
forests. Forest Ecology and Management 500. https://doi.org/10.1016/j.foreco.2021.119629
Bolstad, P. V., Elliott, K.J., Miniat, C.F. 2018. Forests, shrubs, and
terrain: top-down and bottom-up controls on forest structure. Ecosphere
9. https://doi.org/10.1002/ecs2.2185
Caparros-Santiago, J.A.,
Rodríguez-Galiano, V.F. 2020. Vegetation phenology from
satellite imagery: The case of the iberian peninsula and balearic islands
(2001-2017). Revista de Teledeteccion 2020: 25-36.
Chen, C., Park, T., Wang, X., Piao, S., Xu, B., Chaturvedi, R.K., Fuchs,
R., et al. 2019. China and India lead in greening of the world through land-use
management. Nature Sustainability 2: 122-129. https://doi.org/10.1038/s41893-019-0220-7
Cortés, J., Mahecha, M.D., Reichstein, M., Myneni, R.B., Chen, C.,
Brenning, A. 2021. Where Are Global Vegetation Greening and Browning Trends
Significant? Geophysical Research Letters 48. https://doi.org/10.1029/2020GL091496
Dalezios, N.R., Blanta, A., Spyropoulos, N. V., Tarquis, A.M. 2014. Risk identification of agricultural drought for sustainable
Agroecosystems. Natural Hazards and Earth System Sciences 14: 2435-2448.
https://doi.org/10.5194/nhess-14-2435-2014
Díaz, M., Sánchez-Mejía, T.,
Morán-López, T. 2021. Long-Term Tree Regeneration of
Fragmented Agroforestry Systems Under Varying Climatic Conditions. Frontiers
in Ecology and Evolution 9. https://doi.org/10.3389/fevo.2021.640143
Durante, P., Martín-Alcón, S.,
Gil-Tena, A., Algeet, N., Tomé, J.L., Recuero, L., Palacios-Orueta, A., et al.
2019. Improving aboveground forest biomass maps: From
high-resolution to national scale. Remote Sensing 11. https://doi.org/10.3390/rs11070795
Ellison, D., Morris, C.E., Locatelli, B., Sheil, D., Cohen, J., Murdiyarso,
D., Gutierrez, V., et al. 2017. Trees, forests and water: Cool insights for a
hot world. Global Environmental Change 43: 51-61. https://doi.org/10.1016/j.gloenvcha.2017.01.002
FAO. 2003. State of the World´s Forests 2003 Management,
conservation and sustainable development of forests. Role of planted trees in
developing countries with low forest cover: findings from six case studies. https://www.fao.org/docrep/pdf/005/y7581e/y7581e02.pdf
FAO. 2020. Global Forest Resources Assessment 2020 – Key findings. Rome.
https://doi.org/10.4060/ca8753en
Ferraz-de-Oliveira, M.I., Azeda, C., Pinto-Correia, T. 2016. Management
of Montados and Dehesas for High Nature Value: an interdisciplinary pathway. Agroforestry
Systems 90: 1-6. https://doi.org/10.1007/s10457-016-9900-8
Franklin, J.F., Spies, T.A., Van Pelt, R., Carey, A.B., Thornburgh, D.A.,
Berg, R., Lindenmayer, D.B., et al. 2002. Disturbances and structural
development of natural forest ecosystems with silvicultural implications, using
Douglas-fir forests as an example. Forest Ecology and Management 155(1-3):
399-423. https://doi.org/10.1016/s0378-1127(01)00575-8
Freitas, S.R., Mello, M.C.S., Cruz, C.B.M. 2005. Relationships
between forest structure and vegetation indices in Atlantic Rainforest. Forest
Ecology and Management 218: 353-362. https://doi.org/10.1016/j.foreco.2005.08.036
Gea-Izquierdo, G., Cañellas, I., Montero, G. 2008. Site index in agroforestry
systems: Age-dependent and age-independent dynamic diameter growth models for Quercus
ilex in Iberian open oak woodlands. Canadian Journal of Forest
Research 38: 101-113. https://doi.org/10.1139/X07-142
González Díaz, P., Ruiz
Benito, P., Astigarraga Urcelay, J., Cruz Alonso, V., Moreno Fernández, D.,
Herrero Méndez, A., Gosálbez Ruiz, J., et al. 2020. Los bosques españoles
como soluciones naturales frente al cambio climático: herramientas de análisis
y modelización. Madrid.
Gorelick, N., Hancher, M., Dixon, M., Ilyushchenko, S., Thau, D., Moore, R.
2017. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote
Sensing of Environment 202: 18-27. https://doi.org/10.1016/j.rse.2017.06.031
Gutiérrez
Hernández, O. 2022. Recent NDVI trends in Andalusia
(southern Spain): the limits of vegetation greening. Boletín de la
Asociación de Geógrafos Españoles, 94. https://doi.org/10.21138/bage.3246
Helman, D., Lensky, I.M., Osem, Y., Rohatyn, S., Rotenberg, E., Yakir, D.
2017. A biophysical approach using water deficit factor for daily estimations
of evapotranspiration and CO2 uptake in Mediterranean environments. Biogeosciences
14: 3909-3926. https://doi.org/10.5194/bg-14-3909-2017
Hernández, L., Alberdi, I.,
Cañellas, I., Barrera, M. 2014. The forest biodiversity of
Andalusia: estimation based on the Spanish NFI data analysis.
Hernández-Silva, D.A., Pulido, M.T., Zuria, I., Gallina Tessaro, S.A., Sánchez-Rojas,
G. 2018. El manejo como herramienta para la conservación y
aprovechamiento de la fauna silvestre: acceso a la sustentabilidad en México. Acta
Universitaria 28: 31-41. https://doi.org/10.15174/au.2018.2171
Herraiz, A.D., Salazar-Zarzosa,
P.C., Mesas, F.J., Arenas-Castro, S., Ruiz-Benito, P., Villar, R. 2023. Modelling aboveground biomass and productivity and the impact of
climate change in Mediterranean forests of South Spain. Agricultural
and Forest Meteorology 337. https://doi.org/10.1016/j.agrformet.2023.109498
Herraiz, A.D., Salazar-Zarzosa,
P., Acosta-Muñoz, C., Hernández-Clemente, R., Villar, R. 2025. Aridity-induced phenological shifts and greening trends in
Mediterranean forest species: Insights from 28 years of Landsat data in
southern Spain. Ecological Indicators 171. https://doi.org/10.1016/j.ecolind.2025.113115
Hijmans, R.J. 2012. Cross-validation of species distribution models:
removing spatial sorting bias and calibration with a null model. Ecology
93: 679-688. https://doi.org/10.1890/11-0826.1
Hoek van Dijke, A.J., Orth, R., Teuling, A.J., Herold, M., Schlerf, M.,
Migliavacca, M., Machwitz, M., et al. 2023. Comparing forest and grassland
drought responses inferred from eddy covariance and Earth observation. Agricultural
and Forest Meteorology 341. https://doi.org/10.1016/j.agrformet.2023.109635
Huang, S., Tang, L., Hupy, J.P., Wang, Y., Shao, G. 2021. A commentary
review on the use of normalized difference vegetation index (NDVI) in the era
of popular remote sensing. Journal of Forestry Research 32. https://doi.org/10.1007/s11676-020-01155-1
Huete, A., Didan, K., Miura, T., Rodriguez, E.P., Gao, X., Ferreira, L.G.
2002. Overview of the radiometric and biophysical performance of the MODIS
vegetation indices. Remote Sensing of Environment 83(1-2): 195-213. https://doi.org/10.1016/S0034-4257(02)00096-2
Jiao, W., Wang, L., McCabe, M.F. 2021. Multi-sensor remote sensing for
drought characterization: current status, opportunities and a roadmap for the
future. Remote Sensing of Environment 256. https://doi.org/10.1016/j.rse.2021.112313
Kendall, M.G. 1975. Rank Correlation Methods. 4th ed. Charles
Griffin, London.
Kim, H.N., Jin, H.Y., Kwak, M.J., Khaine, I., You, H.N., Lee, T.Y.,
Ahn, T.H., et al. 2017. Why does Quercus suber species decline in
Mediterranean areas? Journal of Asia-Pacific Biodiversity 10: 337-341. https://doi.org/10.1016/j.japb.2017.05.004
Kladny, K.R., Milanta, M., Mraz, O., Hufkens, K., Stocker, B.D. 2024.
Enhanced prediction of vegetation responses to extreme drought using deep
learning and Earth observation data. Ecological Informatics 80. https://doi.org/10.1016/j.ecoinf.2024.102474
Kogan, F.N. 1995. Application of vegetation index and brightness
temperature for drought detection. Advances in Space Research 15(11),
91-100. https://doi.org/10.1016/0273-1177(95)00079-T
Laamrani, A., Valeria, O., Bergeron, Y., Fenton, N., Cheng, L.Z., Anyomi, K.
2014. Effects of topography and thickness of organic layer on productivity of
black spruce boreal forests of the canadian clay belt region. Forest Ecology
and Management 330: 144-157. https://doi.org/10.1016/j.foreco.2014.07.013
Li, A., Yin, G., Zhang, Z., Tan, J., Nan, X., Ma, K., Guo, Q. 2018.
Space-air-field integrated biodiversity monitoring based on experimental
station. Biodiversity Science 26: 819-827. https://doi.org/10.17520/biods.2018052
López-Tirado,
J., Hidalgo, P.J. 2016. Predictive modelling of
climax oak trees in southern Spain: insights in a scenario of global change. Plant
Ecology 217: 451-463. https://doi.org/10.1007/s11258-016-0589-6
Mann, H.B. 1945. Nonparametric Tests Against Trend. Econometrica
13, 3 (1945): 245–59. https://doi.org/10.2307/1907187
Mantero, G., Morresi, D., Marzano,
R., Motta, R., Mladenoff, D.J., Garbarino, M. 2020. The
influence of land abandonment on forest disturbance regimes: a global review. Landscape
Ecology 35: 2723-2744. https://doi.org/10.1007/s10980-020-01147-w
Marañón, T., Ibáñez, B.,
Anaya-Romero, M., Muñoz-Rojas, M., Pérez-Ramos, I.M. 2012. Trees
beyond the wood conference proceedings Oak trees and woodlands providing
ecosystem services in Southern Spain.
Moreno, G., Pulido, F.J.
2009. The Functioning, Management and Persistence of Dehesas.
In: Rigueiro-Rodríguez, A. et al. (eds.), Agroforestry in Europe, pp.
127-160. Springer Science + Business Media B.V. https://doi.org/10.1007/978-1-4020-8272-6_7
Moreno, G., Aviron, S., Berg, S.,
Crous-Duran, J., Franca, A., de Jalón, S.G., Hartel, T., et al. 2018. Agroforestry systems of high nature and cultural value in Europe:
provision of commercial goods and other ecosystem services. Agroforestry
Systems 92: 877-891. https://doi.org/10.1007/s10457-017-0126-1
Mutanga, O., Masenyama, A., Sibanda, M. 2023. Spectral
saturation in the remote sensing of high-density vegetation traits: A
systematic review of progress, challenges, and prospects. ISPRS Journal of
Photogrammetry and Remote Sensing 198: 297-309. https://doi.org/10.1016/j.isprsjprs.2023.03.010
Naranjo Ramírez, J. 2003. El
campo andaluz. En: López Ontiveros, A. (coord.), Geografía de Andalucía,
pp. 523-548. Ariel. Barcelona, España.
Nesha, K., Herold, M., De Sy, V., de
Bruin, S., Araza, A., Málaga, N., Gamarra, J.G.P., et al. 2022. Exploring characteristics of national forest inventories for
integration with global space-based forest biomass data. Science of the
Total Environment 850. https://doi.org/10.1016/j.scitotenv.2022.157788
Oksuz, D.P., Aguiar, C.A.S., Tápia, S., Llop, E., Lopes, P., Serrano,
A.R.M., Leal, A.I., et al. 2020. Increasing biodiversity in wood-pastures by
protecting small shrubby patches. Forest Ecology and Management 464. https://doi.org/10.1016/j.foreco.2020.118041
Pardos, M., del Río, M., Pretzsch, H., Jactel, H., Bielak, K., Bravo, F.,
Brazaitis, G. et al. 2021. The greater resilience of mixed forests to drought
mainly depends on their composition: Analysis along a climate gradient across
Europe. Forest Ecology and Management 481. https://doi.org/10.1016/j.foreco.2020.118687
Parra-López, C., Sayadi, S., Garcia-Garcia, G., Ben Abdallah, S.,
Carmona-Torres, C. 2023. Prioritising conservation actions towards the
sustainability of the dehesa by integrating the demands of society. Agricultural
Systems 206. https://doi.org/10.1016/j.agsy.2023.103613
Pulido, F., Picardo, Á., Campos,
P., Carranza, J., Coleto, J.M., Díaz, M., Diéguez, E., et al. 2010. Libro
Verde de la Dehesa. Documento para el debate hacia un Estrategia Ibérica de
gestión. Consejería de Medio Ambiente, Junta de Castilla y León; SECF;
SEEP; AEET y SEO. España.
Rasoanaivo, A., Mekontchou, C.G., Rochon, P., Nolet, P., Maheu, A. 2024. Increased tree water use with the development of a dense understory
layer in a North American hardwood forest. Ecohydrology 17. https://doi.org/10.1002/eco.2648
Rendón-Pérez, M.A.,
Hernández-De la Rosa, P., Velázquez-Martínez, A., Alcántara-Carbajal, J.L.,
Reyes-Hernández, V.J. 2021. Composition, diversity, and
structure of a managed forest in central mexico. Madera y Bosques 27. https://doi.org/10.21829/myb.2021.2712127
Requena-Mullor, J.M., Steiner, A., Keppel-Aleks, G., Ibáñez, I. 2023. Tradeoffs in
forest resilience to satellite-based estimates of water and productivity
losses. Remote Sensing of Environment 285. https://doi.org/10.1016/j.rse.2022.113414
Rouse, J.W., Haas, R.H., Schell, J.A., Deering, D.W. 1973. Monitoring
Vegetation Systems in the Great Plains with ERTS (Earth Resources Technology
Satellite). In:
Third Earth Resources Technology Satellite-1 Symposium. Volume 1: Technical
Presentations, section A, Paper A20. (Work 19740022614). NASA, US
Goverment. https://ntrs.nasa.gov/api/citations/19740022614/downloads/19740022614.pdf
Ruiz-Benito, P.,
Gómez-Aparicio, L., Paquette, A., Messier, C., Kattge, J., Zavala, M.A. 2014. Diversity increases carbon storage and tree productivity in Spanish
forests. Global Ecology and Biogeography 23: 311-322. https://doi.org/10.1111/geb.12126
Ruiz-Benito, P., Ratcliffe, S., Jump, A.S., Gómez-Aparicio, L.,
Madrigal-González, J., Wirth, C., Kändler, G., et al. 2017. Functional
diversity underlies demographic responses to environmental variation in
European forests. Global Ecology and Biogeography 26: 128-141. https://doi.org/10.1111/geb.12515
Ruiz-Peinado, R., Montero, G.,
del Rio, M. 2012. Ecuaciones para la estimación de biomasa de frondosas en
España. Forest Systems 21: 42-52. https://doi.org/10.5424/fs/2112211-02193
Salazar-Zarzosa, P., Diaz
Herraiz, A., Olmo, M., Ruiz-Benito, P., Barrón, V., Bastias, C.C., de la Riva,
E.G., et al. 2021. Linking functional traits with tree growth
and forest productivity in Quercus ilex forests along a climatic
gradient. Science of the Total Environment 786. https://doi.org/10.1016/j.scitotenv.2021.147468
Spasojevic, M., Shprits, Y.Y., Orlova, K. 2015. Global empirical models of
plasmaspheric hiss using Van Allen Probes. Journal of Geophysical Research:
Space Physics 120: 10370-10383. https://doi.org/10.1002/2015JA021803
Stadler, S.J. 2005. Aridity Indexes. In: Oliver, J.E. (Ed.), Encyclopedia
of World Climatology. Encyclopedia of Earth Sciences Series, pp. 89-94.
Springer, Dordrecht. The Netherlands.
Torres, P., Rodes‐blanco, M., Viana‐soto, A., Nieto, H., García,
M. 2021. The role of remote sensing for the assessment and
monitoring of forest health: A systematic evidence synthesis. Forests
12. https://doi.org/10.3390/f12081134
Valladares, F., Matesanz, S.,
Guilhaumon, F., Araújo, M.B., Balaguer, L., Benito-Garzón, M., Cornwell, W., et
al. 2014. The effects of phenotypic plasticity and local
adaptation on forecasts of species range shifts under climate change. Ecology Letters 17: 1351-1364. https://doi.org/10.1111/ele.12348
Vicente-Serrano, S.M.,
Martín-Hernández, N., Camarero, J.J., Gazol, A., Sánchez- Salguero, R.,
Peña-Gallardo, M., El Kenawy, A., et al. 2020. Linking
tree-ring growth and satellite-derived gross primary growth in multiple forest
biomes. Temporal-scale matters. Ecological Indicators. 108. https://doi.org/10.1016/j.ecolind.2019.105753
Vila-Viçosa, C., Arenas-Castro, S., Marcos, B., Honrado, J., García, C., Vázquez,
F.M., Almeida, R., Gonçalves J. 2020. Combining Remote
Sensing and Climate Data in Species Distribution Models to improve the
conservation of the Iberian white oaks (Quercus L.). ISPRS
International Journal of Geo-Information, 9:735. https://doi.org/10.3390/ijgi9120735
Vio, R.,
Wamsteker, W. 2001. Limits of the
Cross-Correlation Function in the Analysis of Short Time Series. PASP 113,
86-97. https://doi.org/10.1086/317967
Wang, Q.,
Tenhunen, J.D. 2004. Vegetation mapping with
multitemporal NDVI in North Eastern China Transect (NECT). International
Journal of Applied Earth Observation and Geoinformation 6: 17-31. https://doi.org/10.1016/j.jag.2004.07.002
Wang, S., Zhou, L., Chen, J., Ju, W., Feng, X., Wu, W. 2011.
Relationships between net primary productivity and stand age for several forest
types and their influence on China’s carbon balance. Journal of
Environmental Management 92: 1651-1662. https://doi.org/10.1016/j.jenvman.2011.01.024
Wang, Y., Sun, Y., Cao, X., Wang, Y., Zhang, W., Cheng, X. 2023. A
review of regional and Global scale Land Use/Land Cover (LULC) mapping products
generated from satellite remote sensing. ISPRS Journal of Photogrammetry and
Remote Sensing 206: 311-334. https://doi.org/10.1016/j.isprsjprs.2023.11.014
Young, D.J.N., Estes, B.L., Gross, S., Wuenschel, A., Restaino, C.,
Meyer, M.D. 2023. Effectiveness of forest density reduction treatments for
increasing drought resistance of ponderosa pine growth. Ecological
Applications 33. https://doi.org/10.1002/eap.2854
Zellou, B., El Moçayd, N., EL Houcine, B. 2023. Review
article: Towards improved drought prediction in the Mediterranean region -
Modeling approaches and future directions. Natural Hazards and Earth System
Sciences 23: 3543-3583. https://doi.org/10.5194/nhess-23-3543-2023
Anexo / Appendix
Figuras A1-A7 / Figures A1-A7
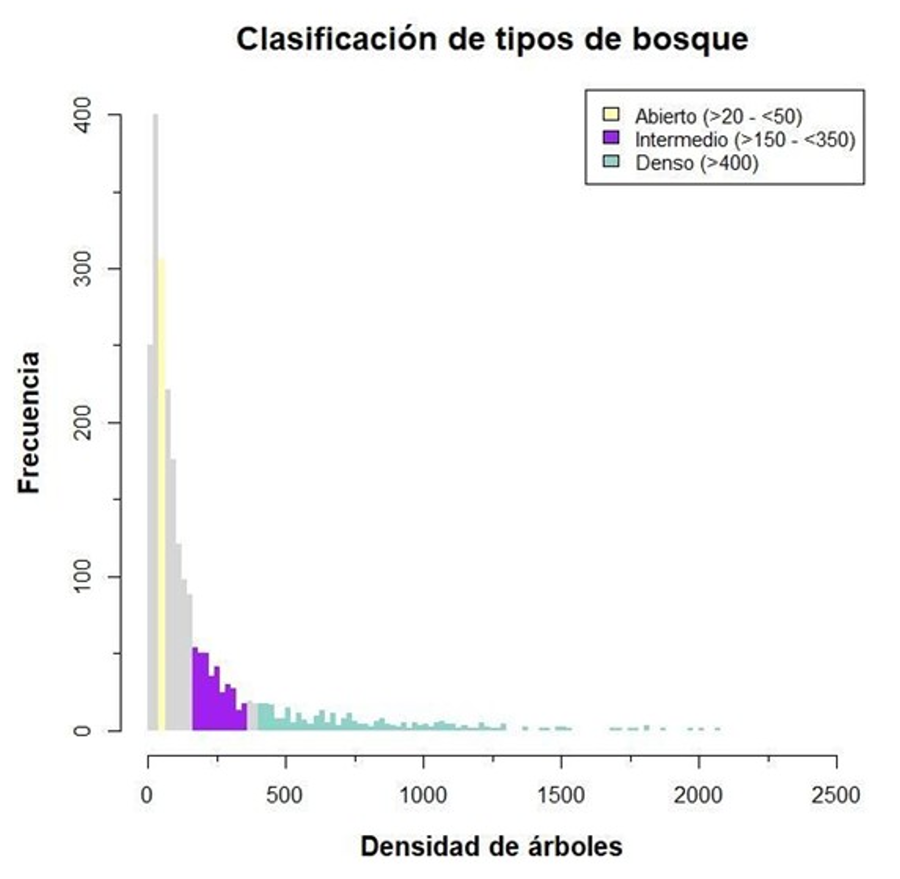
Figura A1. Clasificación de los tipos de
bosque de Q. ilex en base a la densidad de arbolado.
Figure A1. Classification of Q. ilex forest type based on tree density.
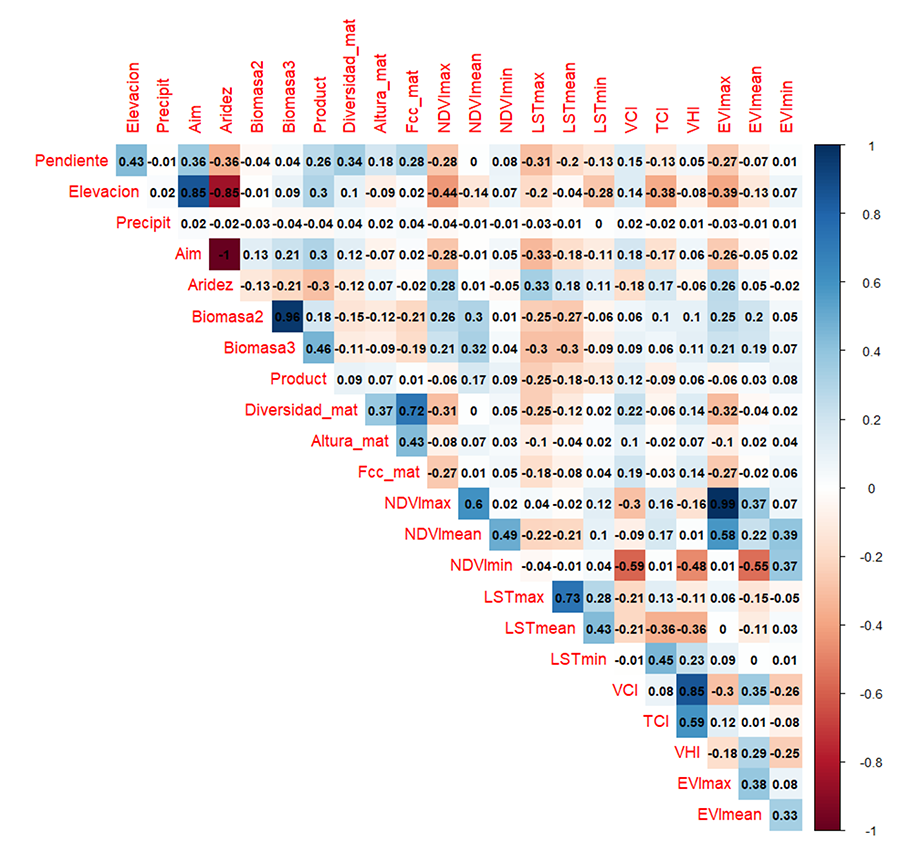
Figura A2. Coeficiente
de correlación de Spearman de todas las variables. AIm: Índice de
Aridez de Martonne; NDVImax: Índice de Diferencia Normalizada de la
Vegetación (máximo anual); NDVImedio: Índice de Diferencia
Normalizada de la Vegetación (media anual); NDVImin: Índice de
Diferencia Normalizada de la Vegetación (mínimo anual); LSTmax:
Temperatura Superficial (máxima anual); LSTmean: Temperatura
superficial (media anual); LSTmin: Temperatura Superficial (mínima
anual); VCI: Índice de las Condiciones de la Vegetación; TCI: Índice de las
Condiciones de Temperatura; VHI: Índice de Salud de la Vegetación; EVImax:
Índice de Vegetación Mejorado (máximo anual); EVImedio: Índice de
Vegetación Mejorado (media anual); ; EVImin: Índice de
Vegetación Mejorado (mínimo anual).
Figure A2. Spearman rank correlation for the variables. AIm:
Martonne Aridity Index; NDVImax: Normalized Difference Vegetation
Index (annual maximum); NDVImedio: Normalized Difference Vegetation
Index (annual mean); NDVImin: Normalized Difference Vegetation Index
(annual minimum); LSTmax: Land Surface Temperature (annual maximum);
LSTmean: Land Surface Temperature (annual mean); LSTmin:
Land Surface Temperature (annual minimum); VCI: Vegetation Condition Index;
TCI: Temperature Condition Index; VHI: Vegetation Health Index. EVImax:
Enhanced Vegetation Index (annual maximum); EVImedio: Enhanced
Vegetation Index (annual mean); EVImin: Enhanced Vegetation
Index (annual minimum); VHI: Vegetation Health Index.
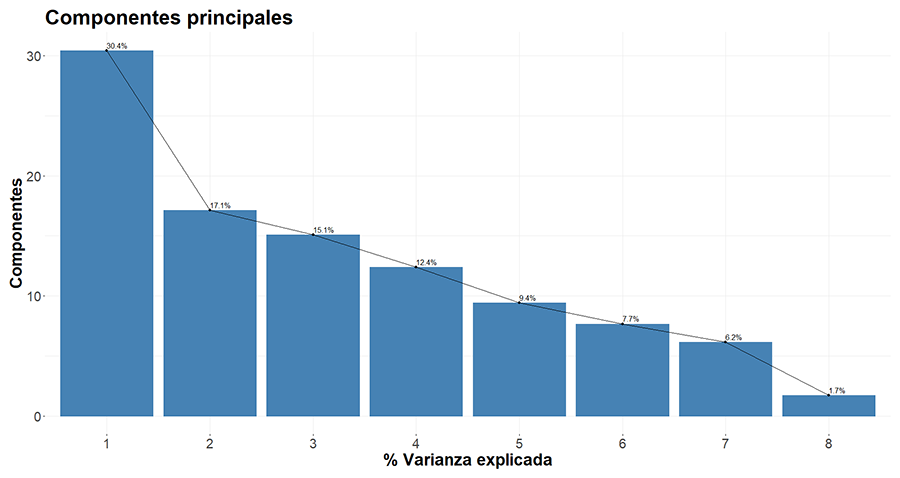
Figura A3. Porcentaje explicado de los
componentes principales del análisis PCA realizado.
Figure A3. Explained percentage of the principal components from the performed
PCA analysis.
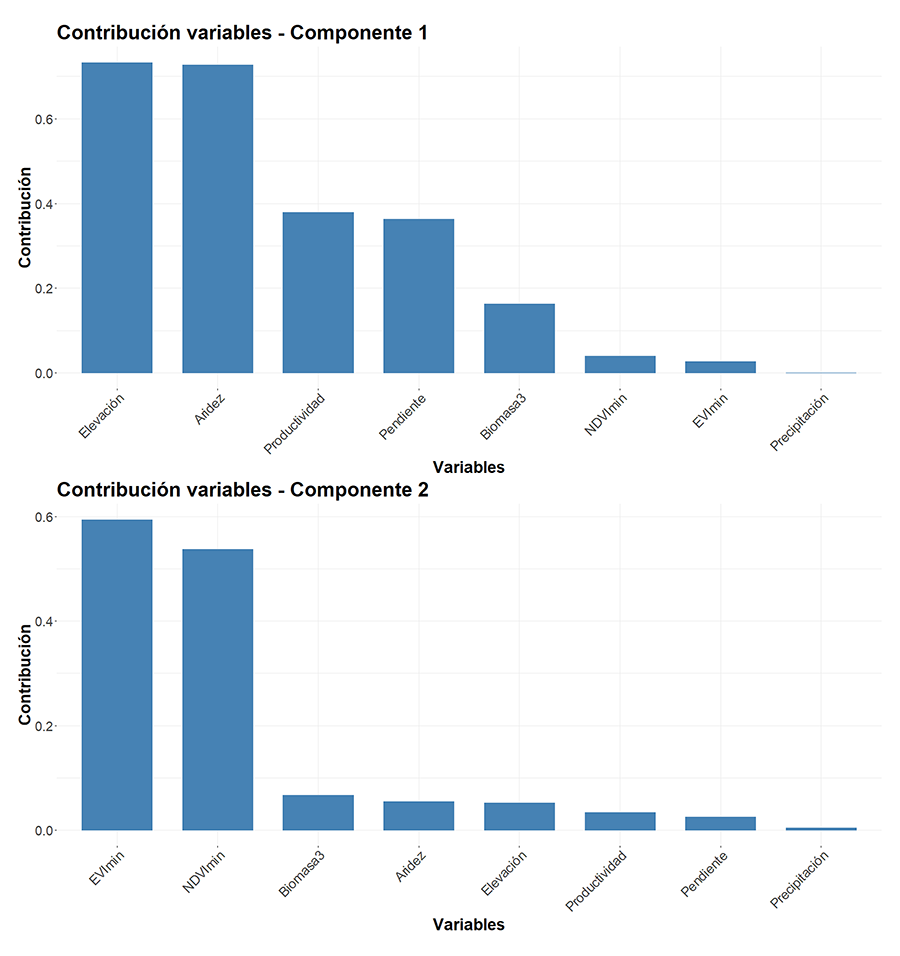
Figura A4. Contribución de las variables
a los dos principales componentes del análisis.
Figure A4. Contribution of the variables to
the first two principal components of the analysis.
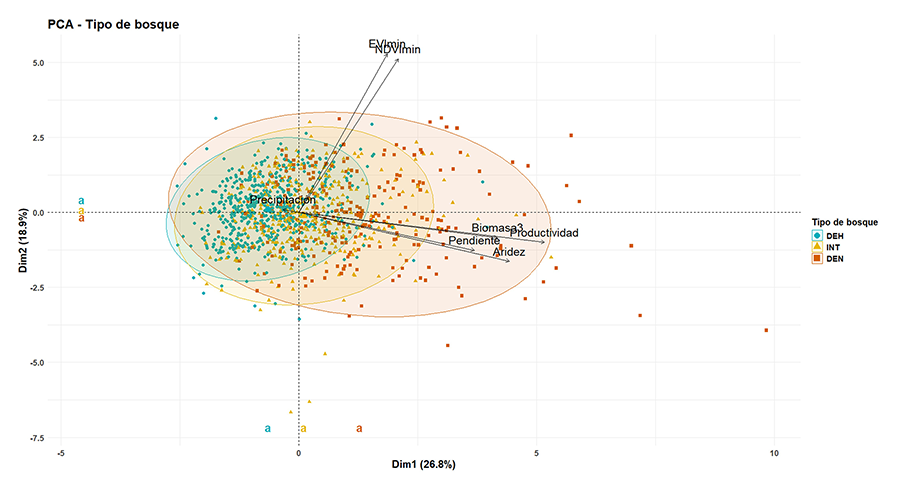
Figura A5. Análisis de componentes principales
de las variables seleccionadas para la diferenciación de tipos de bosque.
Diferentes letras indican una diferencia estadística (P<0.05) entre grupos.
Figure A5. Principal
components analysis (PCA) of the selected variables for forest type
differentiation. Different letters indicate a statistical difference
(P<0.05) between groups.
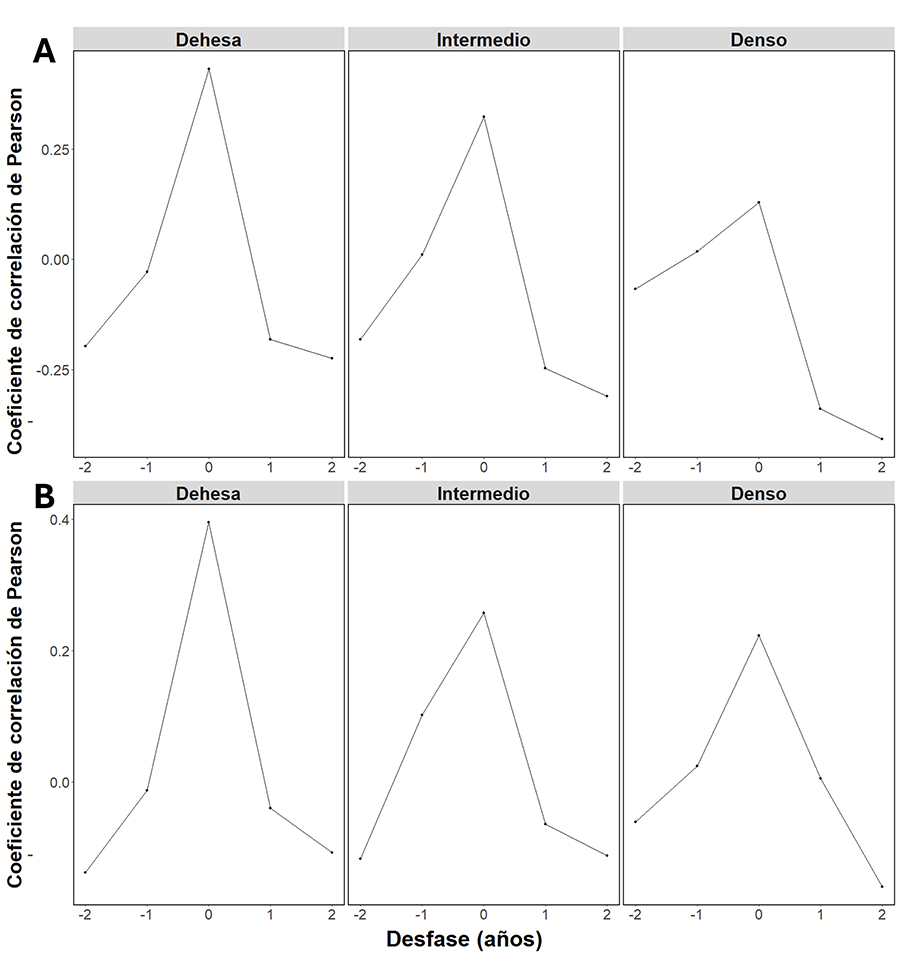
Figura A6. Análisis de correlación
cruzada del NDVImáx (A) y EVImáx (B) con respecto a la
precipitación por tipo de bosque.
Figure A6. Cross-correlation analysis of maximum of NDVI and maximum of EVI
with respect to precipitation by forest type.
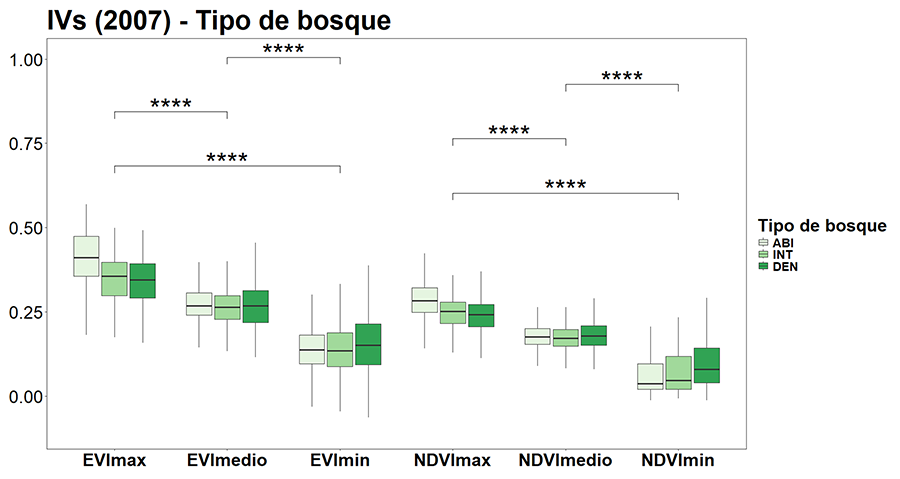
Figura A7.
Comparación de las métricas (máximos, mínimos y media) de los IVs. Límite
inferior de la caja como primer cuartil (Q1), límite superior como tercer
cuartil (Q3) y línea divisoria como mediana o segundo cuartil (Q2). Las barras
indican la desviación estándar. Los asteriscos (***) indican una diferencia
estadística (P<0.05) entre grupos.
Figure A7. Comparison of IVs metrics (maximums, minimums and mean). Lower
boundary of the box as first quartile (Q1), upper boundary as third quartile
(Q3) and dividing line as median or second quartile (Q2). The whiskers indicate
the standard deviation. The asterisks (****) indicate a statistical difference
(P<0.05) between groups.
Tablas A1-A4 / Tables
A1-A4
Tabla A1. Aportación
de las variables del resto de los GLMs para los tres tipos de bosque (ABI,
abierto; INT, intermedio; DEN, denso). Grado de significancia representado por:
*** (p-value <0.001), ** (0.001 < p-value < 0.01), **
(0.001 < p-value < 0.01), * (0.01 < p-value < 0.05). NDVImedio:
Índice de Diferencia Normalizada de la Vegetación (media anual); EVImedio:
Índice de Vegetación Mejorado (media anual); NDVImin: Índice de
Diferencia Normalizada de la Vegetación (mínimo anual); EVImín:
Índice de Vegetación Mejorado (mínimo anual).
Table A1.
Contribution of the variables from the rest of
the GLMs for the three forest types (ABI, open; INT, intermediate; DEN, dense).
Degree of significance represented by: *** (p-value < 0.001), **
(0.001 < p-value < 0.01), * (0.01 < p-value < 0.05).
NDVImedio: Normalized Difference Vegetation Index (annual mean);
EVImedio: Enhanced Vegetation Index (annual mean); NDVImin:
Normalized Difference Vegetation Index (annual minimum); EVImin:
Enhanced Vegetation Index (annual minimum); VHI: Vegetation Health Index.
Tabla A2.
Métricas de rendimiento de los modelos lineales generalizados (GLMs)
con distinto índice espectral como variable respuesta para el ABI. NDVImax:
Índice de Diferencia Normalizada de la Vegetación (máximo anual); EVImax:
Índice de Vegetación Mejorado (máximo anual); NDVImedio: Índice de
Diferencia Normalizada de la Vegetación (media anual); EVImedio:
Índice de Vegetación Mejorado (media anual); NDVImin: Índice de
Diferencia Normalizada de la Vegetación (mínimo anual); EVImin:
Índice de Vegetación Mejorado (mínimo anual); VHI: Índice de salud de la
vegetación.
Table A2.
Generalized linear model (GLMs) performance
metrics from each spectral index as response variable for ABI. NDVImax:
Normalized Difference Vegetation Index (annual maximum); EVImax:
Enhanced Vegetation Index (annual maximum); NDVImedio: Normalized
Difference Vegetation Index (annual mean); EVImedio: Enhanced
Vegetation Index (annual mean); NDVImin: Normalized Difference
Vegetation Index (annual minimum); EVImin: Enhanced Vegetation Index
(annual minimum); VHI: Vegetation Health Index.
Tabla A3. Métricas
de rendimiento de los modelos lineales generalizados (GLMs) con distinto índice
espectral como variable respuesta para el INT. NDVImax: Índice de
Diferencia Normalizada de la Vegetación (máximo anual); EVImax:
Índice de Vegetación Mejorado (máximo anual); NDVImedio: Índice de
Diferencia Normalizada de la Vegetación (media anual), EVImedio:
Índice de Vegetación Mejorado (media anual); NDVImin: Índice de
Diferencia Normalizada de la Vegetación (mínimo anual); EVImin:
Índice de Vegetación Mejorado (mínimo anual); VHI: Índice de salud de la
vegetación.
Table A3.
Generalized linear model (GLMs) performance
metrics from each spectral index as response variable for INT. NDVImax:
Normalized Difference Vegetation Index (annual maximum); EVImax:
Enhanced Vegetation Index (annual maximum); NDVImedio: Normalized
Difference Vegetation Index (annual mean); EVImedio: Enhanced
Vegetation Index (annual mean); NDVImin: Normalized Difference
Vegetation Index (annual minimum); EVImin: Enhanced Vegetation Index
(annual minimum); VHI: Vegetation Health Index.
Tabla A4.
Métricas de rendimiento de los modelos lineales generalizados (GLMs)
con distinto índice espectral como variable respuesta para el DEN. NDVImax:
Índice de Diferencia Normalizada de la Vegetación (máximo anual); EVImax:
Índice de Vegetación Mejorado (máximo anual); NDVImedio: Índice de
Diferencia Normalizada de la Vegetación (media anual), EVImedio:
Índice de Vegetación Mejorado (media anual); NDVImin: Índice de
Diferencia Normalizada de la Vegetación (mínimo anual); EVImin:
Índice de Vegetación Mejorado (mínimo anual); VHI: Índice de salud de la
vegetación.
Table A4.
Generalized linear model (GLMs) performance
metrics from each spectral index as response variable for DEN. NDVImax:
Normalized Difference Vegetation Index (annual maximum); EVImax:
Enhanced Vegetation Index (annual maximum); NDVImedio: Normalized
Difference Vegetation Index (annual mean); EVImedio: Enhanced
Vegetation Index (annual mean); NDVImin: Normalized Difference
Vegetation Index (annual minimum); EVImin: Enhanced Vegetation Index
(annual minimum); VHI: Vegetation Health Index.