Introducción
Los herbarios son infraestructuras científicas donde se
conservan colecciones vegetales en forma de pliegos, que son muestras
deshidratadas, prensadas, etiquetadas y custodiadas en una instalación diseñada
para tal fin (Paton et al. 2020). Además de la
conservación de pliegos, los herbarios desarrollan otras funciones como la
difusión de la información, la restauración de las muestras y la promoción de
la investigación en taxonomía, sistemática, ecología y medio ambiente (Delnatte 2015). Un pliego de herbario aporta
información geográfica, ecológica, fenológica, fenotípica e histórica,
contenida tanto en la propia muestra vegetal como en los datos de la etiqueta y
su equivalente en los metadatos almacenados digitalmente. Los datos de fecha de
recolección, localización precisa, hábitat, estado fenológico, morfología o
interacciones bióticas son particularmente útiles para estudios de sistemática
y biogeografía (Greve et al. 2016; Lang
et al. 2019). Además, el pliego es el testimonio de una planta recogida en
un lugar y un momento, lo que permite estudiar posibles cambios adaptativos o
biogeográficos, una información importante en factores de cambio global como
las invasiones biológicas, el cambio climático, la destrucción de hábitats y la
contaminación (Funk 2003; Lang et al.
2019).
En el caso de las plantas invasoras, el estudio de los
pliegos disponibles en herbarios permite establecer una cronología del proceso
de invasión y su dinámica poblacional desde los primeros registros en un
territorio hasta el momento actual, y predecir su expansión en dicho territorio
(Exposito-Alonso et al. 2018; Magona et al. 2018). La reconstrucción del proceso de
invasión permite identificar las características de la especie que explican su
éxito como invasora, mejorar el conocimiento sobre su biología y localizar
factores ambientales que expliquen el comportamiento invasor. Esta información
tiene aplicación directa en el diseño de acciones de prevención y gestión (Vilà et al. 2011; Aikio et al. 2012;
Dodd et al. 2016). La información contenida en los
pliegos también puede establecer el origen de una invasora, como en el caso de Arundo
donax (Poaceae), una especie invasora en Europa, Oceanía, África y América,
donde la secuenciación del ADN a partir de ejemplares recolectados en campo y
de pliegos de herbario permitió precisar el origen de las poblaciones europeas
(Hardion et al. 2014).
De los pliegos también se puede obtener información acerca
de las interacciones de las plantas con parásitos, patógenos o herbívoros (Lang et al. 2019). Al igual que ocurre con los procesos de
invasión, la presencia de parásitos en pliegos permite datar y localizar el
origen y documentar cambios en la dinámica de la interacción (Meineke y Davies 2018). De esta forma se ha podido
determinar el origen de herbívoros que atacan a plantas invasoras, como el caso
del lepidóptero Cameraria ohridella (Deschka and Dimić, 1986)
(Gracillariidae), que se alimenta de las hojas de Aesculus hippocastanum
L. (Sapindaceae). La identidad, el origen y la dinámica temporal de la especie
invasora pudo evaluarse a partir de las muestras de herbario y la secuenciación
de su ADN (Lees et al. 2011).
Cortaderia selloana (Schult. & Schult.f.) Asch. & Graebn (Poaceae) es
una gramínea herbácea perenne de gran porte, que florece formando grandes
panículas en forma de plumero que pueden superar cuatro metros de altura (Harradine 1991; Herrera y
Campos 2006; Lázaro-Lobo et al. 2024b). Esta
floración característica le confiere un gran valor ornamental, motivo por el
que se introdujo en diferentes lugares como Estados Unidos, Europa, Sudáfrica o
Australia desde su área nativa en Argentina, Uruguay, Chile, Bolivia, Paraguay
y Brasil (Astegiano 1995; Lázaro-Lobo et al. 2024b). Las inflorescencias
secas y la planta viva para uso ornamental se introdujeron en Europa y
popularizaron por su vistosidad en los siglos XIX y XX (Lambrinos
2001; Domingues y Freitas 2002).
Actualmente, C.
selloana se encuentra naturalizada en gran parte del sur de Europa,
incluyendo las áreas costeras peninsulares, y presenta un comportamiento
manifiestamente invasor con tendencia expansiva (Sanz
Elorza et al. 2004; Herrera y Campos 2006; Tarabon et al. 2018; Pardo-Primoy
y Fagúndez 2019). En España, el primer registro de la especie como planta
espontánea no cultivada data de 1953 en la bahía de Santander, Cantabria (Herrera y Campos 2006). En Galicia las primeras
citas son de principios de los años 80 (González 1988);
en otras comunidades, no hay referencias claras o son más tardías (Sanz Elorza et al. 2004).
En las áreas
invadidas, Cortaderia selloana se reconoce como una de las especies
exóticas invasoras más dañinas presentes en España, por impacto en el paisaje,
la salud humana y la biodiversidad (Fagúndez y
Barrada 2007). La especie invade y modifica medios naturales como
matorrales templados, zonas fluviales, dunas y marismas (Sanz Elorza et al. 2004; Herrera
y Campos 2006). Sin embargo, generalmente se desarrolla en hábitats
antrópicos, donde puede formar comunidades monoespecíficas, asociadas a
infraestructuras viarias como carreteras y autovías, zonas industriales o
periurbanas (Fagúndez y Lema 2019; Pardo-Primoy y Fagúndez 2019). También
colonizan cultivos, prados transformados o repoblaciones, en sistemas ruderales
o sometidos a perturbaciones como incendios o procesos erosivos (Fagúndez y Lema 2019; Pardo-Primoy y Fagúndez 2019).
La gran capacidad de dispersión y colonización de C.
selloana se explica por el éxito de su reproducción sexual. La especie
presenta ginodioecia, con individuos hermafroditas que forman grandes
cantidades de polen y, raramente, frutos (Connor 1973)
e individuos femeninos que producen grandes cantidades de frutos en forma de
cariópside que se dispersan por el viento asistidos por el lema, que es
persistente y peloso (Knowles y Ecroyd 1985; Herrera y Campos 2006). Recientemente se ha
descrito Spanolepis selloanae Gagné, un díptero cecidómido asociado a C.
selloana que evita la formación de semillas y que muestra altos valores de
prevalencia (Fagúndez et al. 2021). Este díptero se
encuentra en la actualidad al menos en el norte y noroeste de España, y toda la
costa de Portugal (Fagúndez et al. 2021; Roldão Almeida et al. 2023; Lázaro-Lobo et al. 2024b), pero se desconoce su
origen y su distribución a escala global.
En este estudio se analizó el material depositado en los
herbarios ibéricos para la identificación del proceso de introducción e
invasión de C. selloana. La información contenida en el etiquetado y el
propio pliego puede ayudar a determinar posibles cambios o momentos clave en el
proceso de invasión. Por esta razón, es particularmente importante la
distribución espacial y temporal de los pliegos, así como la información sobre
el hábitat donde se recogió la especie. Además, se exploró la presencia del
cecidómido S. selloanae asociada a las inflorescencias, para determinar
si el proceso de colonización del díptero se produjo en paralelo al de la
planta hospedadora o en un evento independiente.
Es necesario evaluar el papel de los herbarios en el estudio
de las plantas, porque son infraestructuras que se encuentran en un momento
crítico a nivel global (Crisci et al. 2020). En este
caso de estudio se evalúan las necesidades específicas de gestión de los
pliegos de las plantas invasoras, una herramienta con un gran potencial para
mejorar el conocimiento de las dinámicas de naturalización, expansión y
colonización o de las asociaciones bióticas con organismos que pueden ser
potenciales agentes de control biológico.
Material y métodos
El protocolo de intercambio de material entre herbarios
establece el uso público con fines científicos de las colecciones, incluidos
los intercambios entre instituciones. Por esta razón, las consultas y
solicitudes se realizaron a través del herbario de la Universidade de Santiago
de Compostela o herbario SANT según el acrónimo asignado y recogido en el
código internacional Index Herbariorum (https://sweetgum.nybg.org/science/ih/).
Se solicitó información a 36 herbarios ibéricos y, en los que se confirmó la
disponibilidad de pliegos de C. selloana, se cursó la solicitud de
intercambio correspondiente según los modelos proporcionados por cada herbario.
Dos herbarios se consultaron en persona (SANT y SEV) y el resto envió su
material a las instalaciones del herbario SANT, donde se custodiaron los
pliegos hasta completar el estudio. A su recepción, los pliegos se congelaron
durante al menos dos semanas. Se tomaron fotografías de todos los pliegos,
incluyendo detalles de los caracteres clave de las espiguillas. Para cada
pliego se confirmó la identificación, se determinó el sexo y el estado
fenológico de la inflorescencia. En los pliegos en que se constató la presencia
de S. selloanae, se realizó un recuento de larvas sobre una muestra
aleatoria de 50 flores, para calcular su prevalencia (Fagúndez
et al. 2021).
A cada pliego se le asignó un identificador, formado por el
acrónimo del herbario de procedencia y el número de pliego. Se creó una base de
datos con las medidas obtenidas de la observación de la muestra y de la
información incluida en la etiqueta. Las variables registradas y las categorías
asignadas se recogen en la Tabla 1.
El análisis de las
interacciones entre las variables estudiadas (Tabla 1)
se realizó mediante la construcción de tablas de contingencia para cada par de
variables y el contraste de χ2, excepto en los casos en los que
existían valores bajos o ceros en las frecuencias de algunas combinaciones.
Para la realización de la cartografía se utilizó el programa QGIS en su versión
3.10. Las pruebas estadísticas se realizaron con el paquete “gmodels”
implementado en R (Warnes et al. 2024).
Tabla 1. Variables estudiadas en los pliegos de Cortaderia
selloana de herbarios ibéricos.
Table 1. Studied variables of the Cortaderia
selloana iberian herbarium data.
Resultados
De las solicitudes de información realizadas a 36 herbarios,
se obtuvo respuesta de 29. Ocho herbarios manifestaron no tener ningún pliego
de C. selloana en su colección. A los herbarios que contaban con pliegos
de la especie, se les envió la solicitud oficial a través del herbario SANT. Se
recibieron o estudiaron un total de 159 pliegos de C. selloana de 21
herbarios ibéricos (Tabla 2). Cuatro pliegos
provenientes de dos herbarios presentaban un error en la identificación; dos se
identificaron como Cenchrus setaceus (Forssk.) Morrone y dos como Tripidium
ravennae (L.) Scholz. Se adjuntó la correspondiente etiqueta de revisión al
pliego antes de su devolución. Tres pliegos de tres herbarios distintos se
descartaron por provenir de fuera de la península ibérica, dos se recogieron en
Brasil y uno en las Islas Canarias. Otros dos pliegos con un único código
contenían material de más de una planta y se trataron como muestras
independientes en los análisis. Por tanto, los siguientes resultados se
refieren a un total de 155 pliegos de C. selloana recogidos en el
territorio de la península ibérica (Tabla 2; Fig. 1 y Fig. 2).
Tabla 2.
Herbarios consultados en este estudio.
Table 2. Consulted herbaria on this study.
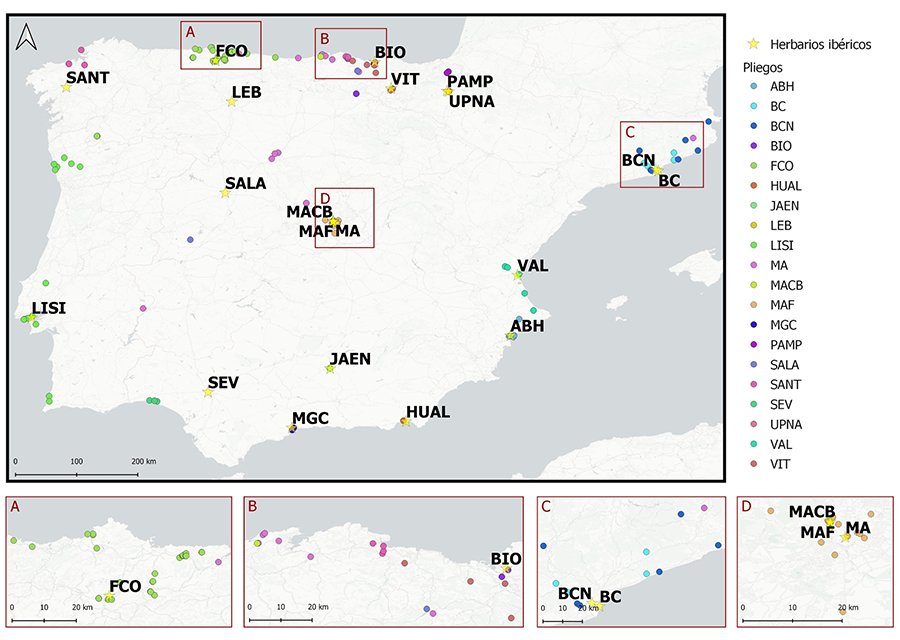
Figura 1.
Localización de los pliegos de Cortaderia selloana y su herbario de
procedencia. Se observa cómo, de forma general, los pliegos se agrupan en torno
a su herbario de origen. (A) Los pliegos del herbario FCO (verde) se
agrupan en torno al herbario, a lo largo de Asturias. (B) Pliegos del
herbario MA (rosa), BIO (morado) y VIT (marrón) distribuidos por el norte
peninsular. (C) Pliegos del herbario MA (rosa), BCN (azul oscuro) y BC
(celeste) distribuidos por Cataluña. (D) Pliegos del herbario MAF (naranja)
distribuidos por el centro peninsular.
Figure 1. Cortaderia selloana herbarium data locations and their herbarium of origin. It can be
appreciated how, mostly, herbarium data are collected close to the herbarium of
origin. (A) FCO
herbarium data (green) grouped around the herbarium, located in Asturias. (B) MA, BIO and VIT herbarium data (pink,
purple, brown) located along the north of the Iberian Peninsula. (C) MA, BCN and BC herbarium data (pink,
dark blue, light blue) located along the north-east of the Iberian Peninsula in
Catalonia. (D) MAF
herbarium data (orange) located in the centre of the Iberian Peninsula.
![Distribución por década de los pliegos de Cortaderia selloana según el carácter invasor [cultivado (rosa), naturalizado (verde) o indeterminado (azul)] y el sexo [femenino (triángulo), hermafrodita (estrella) o indeterminado (cuadrado)].](https://www.revistaecosistemas.net/index.php/ecosistemas/article/download/3014/version/2642/1984/17545/3014-Figura2_web.png)
Figura 2. Distribución por década de los
pliegos de Cortaderia selloana según el carácter invasor [cultivado
(rosa), naturalizado (verde) o indeterminado (azul)] y el sexo [femenino
(triángulo), hermafrodita (estrella) o indeterminado (cuadrado)].
Figure 2.
Distribution of Cortaderia selloana herbarium sheets per decade according to their invasiveness
[cultivated (pink), naturalised (green) or indeterminate (blue)] and sex
[female (triangle), hermaphrodite (star) or indeterminate (square)].
Los pliegos estudiados se recogieron en un periodo que
abarca más de 100 años. El pliego más antiguo de C. selloana procedente
de la península ibérica es un ejemplar femenino recogido en San Martí Sapresa
(Girona) en 1908 y que se conserva en el herbario BCN. La etiqueta original del
pliego aporta poca información, pero incluye una abreviatura (cult.) que parece
indicar un origen de cultivo. Únicamente siete pliegos (4.5 %) son
anteriores a 1980, aunque otros tres que no pudieron datarse son probablemente
anteriores a esta fecha. Otros 30 pliegos se recogieron en las dos últimas
décadas del siglo XX, el resto se recogieron a partir del año 2000. Tres
pliegos (1.9 %) incluían material exclusivamente vegetativo, por lo que no
se determinó su sexo. De los restantes, 107 se identificaron como femeninos (69.0 %)
y 45 como hermafroditas (29.0 %). Ningún pliego recogía esta información
en su etiqueta. Para la fenología de las plantas en flor, de los 152 pliegos en
estadio reproductivo (femeninos o hermafroditas), 64 se categorizaron como
floración temprana (42.1 %), 64 como floración tardía (42.1 %) y en
24 pliegos las inflorescencias se encontraban vacías (15.8 %). Las pruebas
chi-cuadrado mostraron una asociación estadísticamente significativa entre las
variables Sexo y Fenología (χ2 = 16.521; g.l. = 2; p = 0.0002),
con mayor frecuencia de lo esperado de pliegos femeninos con floración tardía
(56) y menor en hermafroditas (8).
En 114 pliegos (73.5 %) se establece de forma clara el
carácter espontáneo de la planta en la etiqueta. En otros 26 pliegos (16.8 %)
se indica procedencia de jardín o cultivo. En los 15 pliegos restantes (9.7 %)
no se constata en la etiqueta si el ejemplar recogido se encontraba
naturalizado o cultivado. Sin considerar estos pliegos indeterminados ni los
vegetativos, se observó una asociación entre las variables Sexo y Origen (χ2
= 4.326; g.l. = 1; p = 0.0375), con mayor frecuencia de lo esperado en
pliegos femeninos naturalizados y en pliegos hermafroditas de cultivo.
La distribución geográfica de los pliegos es sólo
parcialmente coincidente con la distribución actual de la especie naturalizada
(Fig. 1). El mayor número de pliegos de C.
selloana se recogieron en el Cantábrico, a lo largo de gran parte de la
costa e interior del País Vasco, Cantabria y Asturias, y en la costa de
Cataluña. En el interior peninsular se obtuvieron pocas muestras, las más
numerosas de la provincia de Madrid, principalmente como cultivadas, y de
Segovia y Valladolid, como naturalizadas. Portugal está poco representado con
pliegos de tres áreas en el entorno de Porto, Lisboa y Algarve, aunque
únicamente se recibieron pliegos de un herbario portugués (LISI).
Se han encontrado larvas de S.
selloanae en 38 de los 155 pliegos analizados (24.5 %), 35
provenientes de Asturias y depositados en el herbario FCO, y tres de Porto y
Braga, en Portugal, depositados en el herbario LISI. Todas las larvas se
localizaron en pliegos femeninos, con una frecuencia de infección del 35.5 %
de los 107 pliegos femeninos totales (Fig. 3).
Seis pliegos infectados se recogieron en 2018 (15.7 %), y el resto en 2021
(84.2 %). Todos los pliegos infectados se identificaron como fenología
tardía, es decir, en fructificación, excepto uno en fenología temprana, que
presenta ovarios no transformados en las flores. La prevalencia media en las
inflorescencias infectadas, medido en porcentaje de flores con larva sobre un
total de 50, fue de un 84.9 %, con una desviación estándar de 8.056, y un
rango entre el 36 % y el 100 %.

Figura 3. Ejemplo de
las distintas imágenes tomadas de los ejemplares recibidos en el herbario SANT
para la realización del estudio. Se tomaron dos fotografías de cada pliego: una
visión general del individuo del pliego (a) y otra en detalle de la
etiqueta del pliego (b). En caso de presencia de Spanolepis selloanae
también se tomaron fotografías de las larvas (c) [en la imagen se pueden
observar la similitud en tamaño entre tres larvas de S. selloanae (izquierda)
en comparación con una cariópside de C. selloana (derecha)]. En algunos
casos no se pudo confirmar la presencia de S. selloanae en las a partir
de los pliegos de herbario debido al deterioro de las muestras (d) [en
la imagen se puede observar una cariópside de C. selloana (izquierda)
junto a una muestra indeterminada (derecha)].
Figure 3.
Example of the variety of images taken of the herbaria specimen received at
SANT herbarium for the present study. Two pictures were taken of each specimen:
a general overview of the herbarium material received (a) and a closeup of the information
associated with said specimen (b). In relation to the presence of Spanolepis
selloanae, pictures of the larvae were also taken (c) [here depicted the similarities in
size between S. selloanae larvae (left) and C.
selloana caryopsis (right)]. In some cases, the presence
of S. selloanae could
not be confirmed due to sample decay (d) [here depicted C. selloana caryopsis (left) opposed to an undetermined sample (right)].
Discusión
En este trabajo se ha demostrado la utilidad de los pliegos
de herbario, revisados físicamente mediante el procedimiento de solicitud y
préstamo entre herbarios, para describir diferentes aspectos de la invasión de C.
selloana en la península ibérica. Esta especie exótica invasora tiene una
dinámica de expansión elevada y se han señalado los múltiples riesgos que
conlleva la colonización de nuevas áreas (Álvarez Arbesú y González Rodríguez
2023; Cires et al. 2022). La representatividad de
la información obtenida de los pliegos es escasa, considerando la amplia
distribución geográfica y la escala temporal de la invasión. Se han estudiado
155 pliegos, en su mayoría distribuidos en el entorno de los herbarios de referencia
(Fig. 2), con áreas extensas invadidas por la
especie apenas representadas teniendo en cuenta la distribución actual de la
especie en la península ibérica (Roldão Almeida et
al. 2020; Lázaro-Lobo et al. 2024b). También
hay periodos clave en la dinámica temporal de la invasión sin apenas
representación como los 37 pliegos anteriores al 2000, un periodo en que se
produjo la mayor expansión de la especie (Sanz Elorza
et al. 2004). Por ejemplo, en Galicia la especie es muy frecuente en las
áreas costeras y también está presente en zonas del interior (Fagúndez y Barrada 2007). Mediante el uso de
imágenes geolocalizadas de acceso libre y observaciones de campo, se estableció
la presencia de la especie en 59 polígonos industriales de Galicia, con una
tasa de colonización de 1,86 nuevos polígonos al año (Pardo-Primoy y Fagúndez 2019). Sin embargo,
sólo se recibieron cinco pliegos de poblaciones naturalizadas de C. selloana
recolectados en la comunidad gallega. En parte, esto se explica porque las
colecciones de organismos como los herbarios tienden a la sobrerrepresentación
de las especies raras, endémicas y amenazadas, mientras que especies comunes o
introducidas se encuentran infrarrepresentadas (James et
al. 2018).
La relevancia de los herbarios como fuente de información
sobre la distribución de las especies ha variado históricamente. La
digitalización ha facilitado el intercambio de la información contenida en los
herbarios, tanto en forma de bases de datos como de imágenes de los pliegos (James et al. 2018; Borges et al.
2020). Sin embargo, nuevas herramientas de ciencia ciudadana han surgido
como alternativa para la toma de datos de presencia y geolocalización de
especies, especialmente para las especies invasoras que pueden variar su área
de expansión en un corto espacio de tiempo. Por ejemplo, la plataforma
iNaturalist (Matheson 2014) muestra 3899 registros de
C. selloana en la península ibérica (a 15 de marzo de 2025), obtenidas
por 1189 observadores y confirmadas por 457 identificadores. Los registros se
incluyeron a partir de septiembre de 2014, aunque la plataforma admite fechas
de observación anteriores al registro. Este tipo de sistemas permite establecer
la distribución actual de forma rápida y accesible y se ha demostrado su
utilidad para la vigilancia y alerta temprana de especies invasoras (César de Sá et al. 2019), involucrando al público
interesado y aumentando su implicación en la problemática de las invasiones
biológicas (Cordeiro et al. 2020). Al contrario de
lo que ocurre con el material de herbario, los registros de plantas invasoras
tienden a registrarse más que las nativas en las plataformas de ciencia
ciudadana (Dimson et al. 2023).
En este estudio se demuestra que los herbarios tienen otras
prestaciones que demuestran su utilidad frente a fuentes de datos alternativas.
Por ejemplo, la identidad taxonómica de diferentes especies próximas requiere
un análisis de detalle sobre pliego de herbario, frente a fotografías,
observaciones u otras fuentes indirectas como aplicaciones para la
identificación que pueden dar lugar a confusión (White et
al. 2023). En el caso de C. selloana, la identificación frente a la
especie próxima C. jubata (Lemoine) Stapf, considerado en una propuesta
alternativa como subespecie de C. selloana, requiere la
evaluación de un conjunto de caracteres de diagnóstico (Testoni y Villamil 2014). Este estudio permitió
descartar la presencia de C. jubata entre las muestras
recolectadas, y confirma que no hay evidencia de la presencia de esta especie
en la península ibérica. Esta segunda subespecie se caracteriza por su
reproducción apomíctica, y aunque no existen citas confirmadas de la especie
naturalizada en Europa, su presencia en otros lugares como California, en
Estados Unidos, demuestra su capacidad invasora y el riesgo que supondría su
introducción en Europa (Stanton y DiTomaso 2004;
Okada et al. 2009).
En la distribución de las diferentes variables de sexo,
fenología y origen de los pliegos estudiados, se observaron diferentes
variaciones y combinaciones de los estados de las plantas. La frecuencia de los
pliegos hermafroditas provenientes de cultivo fue mayor de lo esperado, al
contrario de los femeninos. Este resultado no se corresponde con la afirmación
de que las plantas cultivadas como ornamentales son principalmente femeninas,
por la mayor espectacularidad de sus inflorescencias (Roldão Almeida et al. 2023), aunque el número de
pliegos es bajo y no permite extraer conclusiones generales. Otras asociaciones
observadas, como la relación entre fenología y sexo, puede ser el resultado de
un sesgo en la recolección del material, algo frecuente en un muestreo no
sistemático como el que ha dado origen a la mayor parte de las colecciones
botánicas (Meyer et al. 2016). Los resultados podrían
indicar que las plantas femeninas se recogen más en la dispersión, cuando las
inflorescencias están abiertas y son más visibles, pero las masculinas se
recogen en estadio temprano porque al madurar pierden volumen y son menos vistosas.
El análisis de los pliegos también permite realizar
prospecciones de patógenos, parásitos, y otra fauna asociada que pueda dejar
algún rastro en las plantas. El análisis confirmó la ausencia de S.
selloanae en todos los pliegos anteriores a 2018 en plantas de cualquier
sexo o estado de maduración, lo que permite establecer la llegada reciente del
díptero a la península ibérica, un aspecto que únicamente puede demostrarse con
la observación de detalle de las inflorescencias. Nuestros resultados también
confirman la alta frecuencia y prevalencia del díptero que se ha descrito para
Galicia (Fagúndez et al. 2021), Asturias (Lázaro-Lobo et al. 2024a) y el norte de Portugal (Roldão Almeida et al. 2023). Este resultado es
coherente con un posible evento de introducción y rápida colonización gracias a
la continuidad de las poblaciones de C. selloana ligadas a grandes vías
de comunicación.
Conclusiones
Los pliegos de herbario tienen un gran potencial para el
estudio de las especies exóticas invasoras en aspectos como la caracterización
genética o la presencia de enemigos naturales asociados. Sin embargo, la planta
invasora C. selloana está infrarrepresentada en los herbarios de la península
ibérica, teniendo en cuenta su distribución espacial y temporal, y es posible
que esta situación sea similar en otras plantas invasoras. Por el contrario, la
recogida de datos por medio de plataformas de ciencia ciudadana como iNaturalist
y otros medios de imágenes remotas pueden aportar un gran volumen de datos y
ser muy útiles en la alerta temprana de las invasiones. Frente a esto, la
recolección de material de especies exóticas y su preparación e inclusión en
los herbarios públicos es una actividad que proporciona información
complementaria insustituible. Proponemos el desarrollo de planes específicos
debidamente financiados con botánicos profesionales, para la realización de
prospecciones periódicas y recolecciones sistemáticas de especies introducidas,
naturalizadas e invasoras. El material debe ser representativo de las
diferentes etapas de desarrollo y otros factores como el sexo en el caso de
plantas dioicas, y recoger la información precisa respecto al grado de
naturalidad de la población. Esta valiosa fuente de información es clave para
garantizar la identidad taxonómica, caracterizar las dinámicas de invasión y
otros aspectos como potenciales estudios genéticos o la búsqueda de enemigos
naturales que puedan servir en el futuro como agentes de control biológico.
Contribución de los autores
Teresa Moreno realizó el trabajo experimental. Jaime
Fagúndez diseñó y dirigió el trabajo, y ambos autores contribuyeron a la
elaboración del artículo.
Disponibilidad de datos y código
Los datos son públicos. La información de los pliegos puede
accederse a través de gbif España y repositorios propios.
Financiación, permisos requeridos, potenciales
conflictos de interés y agradecimientos
Los autores/as declaran no tener ningún conflicto de
intereses.
Los autores quieren agradecer a los técnicos y responsables
de los herbarios contactados por su buena disposición y ayuda. En concreto,
agradecemos a M.B. Crespo Villalba (ABH), F.J. Rejos (AH), N. Ibáñez (BC), J.
Vicens Fandos (BCN), I. García Mijangos (BIO), E. Cires (FCO), C. Salazar
Mendías (JAEN), E. Giménez Luque (HUAL), E. Alfaro Saiz (LEB), P.M.R. Arsénio
(LISI), L. Medina (MA), F.J. Cabezas (MACB), J. Pizarro (MAF), J. García
Sánchez (MGC), M. Imas Lecumberri (PAMP), J. Hernández García (SALA), D.G. San
León (SANT), F.J. Salgueiro González (SEV), J. Peralta de Andrés (UPNA), J.
Riera Vicent (VAL) y P. Heras (VIT). Sirva este artículo como un homenaje a los
botánicos y técnicos y las instituciones que han mantenido la actividad de los
herbarios ibéricos a pesar de las dificultades estructurales que deben
afrontar.
Referencias
Aikio, S., Duncan, R.P., Hulme, P.E.
2012. The vulnerability of habitats to plant invasion:
disentangling the roles of propagule pressure, time and sampling effort. Global
Ecology and Biogeography 21: 778–786. https://doi.org/10.1111/j.1466-8238.2011.00711.x
Álvarez
Arbesú, R.L., González Rodríguez, G. 2023. Viabilidad de la semilla de Cortaderia
selloana en el Principado de Asturias. Naturalia Cantabricae 11:
1-9.
Astegiano, M.E., Anton, A.M.,
Connor, H.E. 1995. Sinopsis del género Cortaderia (Poaceae) en
Argentina. Darwiniana: 43-51.
Borges, L.M., Reis, V.C., Izbicki, R. 2020. Schrödinger's phenotypes:
Herbarium specimens show two‐dimensional images are both good and (not so) bad sources of
morphological data. Methods in Ecology and Evolution 11(10): 1296-1308. https://doi.org/10.1111/2041-210x.13450
César de Sá, N., Marchante, H., Marchante, E., Cabral, J.A., Honrado, J.P.,
Vicente, J.R. 2019. Can citizen science data guide the surveillance of invasive
plants? A model-based test with Acacia trees in Portugal. Biological
Invasions, 21(6): 2127-2141. https://doi.org/10.1007/s10530-019-01962-6
Cires, E., Rafael, D.A.,
González-Toral, C., Cuesta, C. 2022. A preliminary assessment
of the genetic structure of the invasive plant Cortaderia selloana
(Poaceae) in the Iberian Peninsula. Biologia, 77(1): 55-60. https://doi.org/10.1007/s11756-021-00922-7
Connor, H. E. 1973. Breeding systems in Cortaderia (Gramineae). Evolution
27: 663–678. https://doi.org/10.1111/j.1558-5646.1973.tb00714.x
Cordeiro, B., Marchante, H.,
Castro, P., Marchante, E. 2020. Does public awareness about
invasive plants pays off? An analysis of knowledge and perceptions of
environmentally aware citizens in Portugal. Biological Invasions 22(7):
2267-2281. https://doi.org/10.1007/s10530-020-02247-z
Crisci, J.V., Katinas, L., Apodaca, M.J., Hoch, P.C. 2020. The end of botany. Trends in Plant Science, 25(12), 1173-1176. https://doi.org/10.1016/j.tplants.2020.09.012
Delnatte, C. 2015. Sobre la utilidad de los herbarios. Boletín de
la AHIM 17: 19-22.
Dimson, M., Berio Fortini, L.,
Tingley, M.W., Gillespie, T.W. 2023. Citizen science can
complement professional invasive plant surveys and improve estimates of
suitable habitat. Diversity and Distributions 29(9): 1141-1156. https://doi.org/10.1111/ddi.13749
Dodd, A.J., McCarthy, M.A., Ainsworth, N., Burgman, M.A. 2016.
Identifying hotspots of alien plant naturalisation in Australia: approaches and
predictions. Biological Invasions 18: 631–645. https://doi.org/10.1007/s10530-015-1035-8
Domingues,
J., Freitas, H. 2002. Acerca de algunas
plantas vasculares invasoras en Portugal continental. Studia botanica
21: 27-35.
Exposito-Alonso, M.,
Becker, C., Schuenemann, V.J., Reiter, E., Setzer, C., Slovak, R., Brachi, B.,
et al. 2018. The rate and potential relevance of new mutations
in a colonizing plant lineage. PLoS Genetics 14: e1007155. https://doi.org/10.1371/journal.pgen.1007155
Fagúndez, J., Barrada, M.
2007. Plantas invasoras de Galicia. Bioloxía, distribución e métodos de
control. Xunta de Galicia.
Fagúndez, J.,
Lema, M. 2019. A competition experiment of an
invasive alien grass and two native species: are functionally similar species
better competitors? Biological Invasions 21(12): 3619-3631. https://doi.org/10.1007/s10530-019-02073-y
Fagúndez, J., Gagné, R.J., Vila, M. 2021. A new gall
midge species (Diptera, Cecidomyiidae) as a potential candidate for biological
control of the invasive plant Cortaderia selloana (Poaceae). Phytoparasitica
49: 229–241. https://doi.org/10.1007/s12600-020-00844-1
Funk, V. 2003. The importance of herbaria. Plant Science Bulletin
49(3): 94-95.
González, E. 1988. Flora
alóctona gallega. Universidade de
Santiago de Compostela, España.
Greve, M., Lykke, A.M., Fagg, C.W., Gereau, R.E., Lewis, G.P., Marchant,
R., Marshall, A.R., et al. 2016. Realising the
potential of herbarium records for conservation biology. South African
Journal of Botany, 105: 317-323. http://dx.doi.org/10.1016/j.sajb.2016.03.017
Hardion, L., Verlaque, R., Saltonstall, K., Leriche,
A., Vila, B. 2014. Origin of the
invasive Arundo donax (Poaceae): a trans-Asian expedition in herbaria. Annals
of Botany, 114: 455-462. https://doi:10.1093/aob/mcu143
Harradine, A.R. 1991. The impact of pampas grasses as weeds in southern
Australia. Plant Protection Quarterly 6(3): 111-115. https://caws.org.nz/PPQ567/PPQ%2006-3%20pp111-115%20Harradine.pdf
Herrera, M., Campos, J.A.
2006. El carrizo de la Pampa (Cortaderia selloana) en Bizkaia. Guía práctica
para su control. Instituto de Estudios Territoriales de Bizcaia y
Diputación Foral de Bizkaia, España.
James, S.A., Soltis, P.S., Belbin, L., Chapman, A.D., Nelson, G., Paul,
D.L., Collins, M. 2018. Herbarium data: Global biodiversity and societal
botanical needs for novel research. Applications in plant sciences
6(2): e1024. https://doi.org/10.1002/aps3.1024
Knowles, B.,
Ecroyd, C. 1985. Species of Cortaderia
(pampas grasses and toetoe) in New Zealand. Forest Research Bulletin,
105 https://doi.org/10.13140/RG.2.2.22061.95209
Lambrinos, J.G. 2001. The expansion history of a sexual
and asexual species of Cortaderia in California, USA. Journal of
Ecology 89: 88–98. https://doi.org/10.1046/j.1365-2745.2001.00524.x
Lang, P.L.M., Willems, F.M., Scheepens, J.F., Burbano, H.A., Bossdorf,
O. 2019. Using herbaria to study global environmental change. New
Phytologist, 221: 110-122. https://doi.org/10.1111/nph.15401
Lázaro-Lobo, A.,
Alonso-Zaldívar, H., Sagrera, S.J.M., del Alba, C.E., Fernández-Pascual, E.,
González-García, V., Jiménez-Alfaro, B. 2024a. Regeneration
niche of Cortaderia selloana in an invaded region: Flower predation,
environmental stress, and transgenerational effects. Plant Stress, 12:
100483. https://doi.org/10.1016/j.stress.2024.100483
Lázaro-Lobo, A., Andrade, B.O., Canavan, K., Ervin, G.N., Essl, F.,
Fernández-Pascual, E., Follak, S., et al. 2024b. Monographs
on invasive plants in Europe N° 8: Cortaderia selloana (Schult. &
Schult. f.) Asch. & Graebn. Botany Letters, 171(4): 383-407. https://doi.org/10.1080/23818107.2024.2367591
Lees, D. C., Lack, H. W., Rougerie, R., Hernandez-Lopez, A., Raus, T.,
Avtzis, N. D., Lopez-Vaamonde, C. 2011. Tracking origins of invasive herbivores
through herbaria and archival DNA: the case of the horse‐chestnut
leaf miner. Frontiers in Ecology and the Environment 9(6):
322-328. https://doi.org/10.1890/100098
Magona, N., Richardson, D.M., Le Roux, J.J., Kritzinger-Klopper, S.,
Wilson, J.R.U. 2018. Even well-studied groups of alien species might be poorly
inventoried: Australian Acacia species in South Africa as a case study.
NeoBiota 39: 1. https://doi.org/10.3897/neobiota.39.23135
Matheson, C.A. 2014. "iNaturalist". Reference Reviews 28
(8): 36-38. https://doi.org/10.1108/RR-07-2014-0203
Meineke, E.K.,
Davies, T.J. 2018. Museum specimens provide novel
insights into changing plant-herbivore interactions. Philosophical
Transactions R. Soc. B 374: 20170393. http://dx.doi.org/10.1098/rstb.2017.0393
Meyer, C., Weigelt, P., Kreft, H. 2016. Multidimensional biases, gaps and
uncertainties in global plant occurrence information. Ecology letters,
19(8): 992-1006. https://doi.org/10.1111/ele.12624
Okada, M., Lyle, M., Jasieniuk, M. 2009. Inferring the introduction
history of the invasive apomictic grass Cortaderia jubata using
microsatellite markers. Diversity and Distributions 15(1): 148-157. https://doi.org/10.1111/j.1472-4642.2008.00530.x
Pardo-Primoy,
D., Fagúndez, J. 2019. Assessment of the
distribution and recent spread of the invasive grass Cortaderia selloana
in Industrial Sites in Galicia, NW Spain. Flora, 259: 151465. https://doi.org/10.1016/j.flora.2019.151465
Paton, A., Antonelli, A., Carine, M., Forzza, R. C., Davies, N.,
Demissew, S., Dickie, J. 2020. Plant and fungal collections: Current status,
future perspectives. Plants, People, Planet, 2(5):
499-514. ttps://doi.org/10.1002/ppp3.10141
Roldão Almeida, M., García,
A., García, S., González, F., Herrero, A., Marchante, H., Nepomuceno, H., et
al. 2020. Estrategia Transnacional de lucha contra Cortaderia selloana en el
Arco Atlántico en el marco del proyecto LIFE STOP Cortaderia. Consejería de
Desarrollo Rural, Ganadería, Pesca, Alimentación y Medio Ambiente, Gobierno de
Cantabria, España. https://lifecoopcortaderia.org/wp-content/uploads/2024/10/ES_Estrategia-Transnacional_STOP-Cortaderia.pdf
Roldão Almeida, M.,
Marchante, E., Marchante, H. 2023. Public perceptions about
the invasive pampas grass, Cortaderia selloana: a case study of
environmentally conscious citizens in Southern Europe. Biological
Invasions, 25(6): 2043-2056. https://doi.org/10.1007/s10530-023-03025-3
Sanz
Elorza, M., Dana, E., Sobrino, E. 2004. Atlas
de las plantas alóctonas invasoras en España. Dirección General para la
Biodiversidad, España. https://www.miteco.gob.es/es/biodiversidad/temas/inventarios-nacionales/inventario-especies-terrestres/inventario-nacional-de-biodiversidad/ieet_flora_vasc_aloct_invas.html
Stanton,
A.E., DiTomaso, J.M. 2004. Growth response of Cortaderia selloana and
Cortaderia jubata (Poaceae) seedlings to temperature, light, and water. Madrono: 312-321.
Tarabon, S., Bertrand, R., Lavoie, C., Vigouroux, T.,
Isselin-Nondedeu, F. 2018. The effects of climate warming and urbanised areas
on the future distribution of Cortaderia selloana, pampas grass, in
France. Weed Research 58: 413–423. https://doi.org/10.1111/wre.12330
Testoni, D., Villamil, C.B. 2014. Estudios
en el género Cortaderia (Poaceae): I. Sistemática y nomenclatura de la
sect. Cortaderia. Darwiniana, nueva serie 2(2): 260-276. https://doi.org/10.14522/darwiniana/2014.22.591
Vilà, M., Espinar, J.L., Hejda, M.,
Hulme, P.E., Jarosık, V., Maron, J.L., Pergl, J., et al. 2011. Ecological impacts of invasive alien plants: a meta-analysis of
their effects on species, communities and ecosystems. Ecology Letters
14: 702–708. https://doi.org/10.1111/j.1461-0248.2011.01628.x
Warnes, G.R., Bolker, B., Lumley, T., Johnson, R.C., Jain, N., Schwartz,
M., Rogers, J. 2024. _gmodels: Various R Programming Tools for Model
Fitting_. R package version 2.19.1. Available at: https://CRAN.R-project.org/package=gmodels
White, E., Soltis, P.S., Soltis, D.E., Guralnick, R. 2023. Quantifying
error in occurrence data: Comparing the data quality of iNaturalist and
digitized herbarium specimen data in flowering plant families of the
southeastern United States. PLoS One 18(12): e0295298. https://doi.org/10.1371/journal.pone.0295298