Introducción
Desde los albores de la Biología Subterránea (Viré 1900; Racovitza 1907; Martel 1908; Jeannel 1926, 1943; Bonet 1931; Vandel 1965; entre otros), el componente faunístico de
los ecosistemas subterráneos, o hipogeos, ha llamado poderosamente la atención
de los científicos, naturalistas y aventureros que osaban adentrarse en estos
ambientes ignotos. De estrambótica apariencia, muchas de estas especies han
sido consideradas por diversos autores, como el mismo Jeannel
(1943), representantes de linajes arcaicos y tildadas de “fósiles
vivientes”, término que, si bien puede ser relevante desde el punto de vista
tanto teórico como práctico (Turner 2019), ha sido
aplicado con un exceso de ligereza en numerosas ocasiones, siendo la que nos
ocupa ejemplo de ello. Hoy sabemos que, por una parte, es más acertado
considerar a muchas de estas especies como relictas, es decir, especies cuyos
parientes epigeos ya no existen (Ribera et al. 2018)
y, por otra, que el grado de troglobiomorfismo (ver Ortuño
2015: 181) no se correlaciona con la edad del taxón (Delić et al. 2016). Además, desde el comienzo de esta
disciplina científica se observó que los organismos que habitaban el medio
hipogeo mostraban diferentes grados de adaptación al mismo, incluso en especies
pertenecientes a un mismo linaje. Ello condujo a una propuesta de clasificación
ecológica que permitía agrupar a los organismos hipogeos en tres grandes
categorías: troglobios (con mayor o menor grado de troglobiomorfía),
troglófilos (actualmente se distingue entre subtroglófilos y eutroglófilos -ver
Sket 2008-) y trogloxenos (o sus homólogos en la fauna
acuática: estigobios, estigófilos y estigoxenos) (Schiner
1854; Racovitza 1907; Jeannel
1926).
A pesar de que ya Racovitza (1907)
reconocía que la fauna subterránea vivía tanto en las cuevas como en las
grietas que se abrían paso en la roca madre (ver Howarth
1983), no fue hasta la década de 1980, con los trabajos de Christian
Juberthie y colaboradores (Juberthie et al. 1980,
1981) y de Shun-Ichi Uéno (Uéno
1980, 1981), que se materializó un cambio de
paradigma respecto a lo que es, desde un punto de vista biológico, el medio
subterráneo. Ambos autores, de forma independiente (y con distinta
repercusión), pusieron de manifiesto que el dominio subterráneo se extendía desde
el medio subterráneo profundo (MSP sensu Juberthie
1983) hasta el horizonte de disgregación de la roca madre, el llamado medio
subterráneo superficial (MSS). El MSS es un hábitat singular que cuenta
con una fauna hipogea propia (ver, por ejemplo, Juberthie
1983; Casale y Rondolini 1983; Ortuño 1996; Fresneda et
al. 1997; Baquero
et al. 2021a, 2021b),
pero en el que también encontramos, debido a su importante condición de ecotono
(Moseley 2010), tanto especies epigeas y endogeas
como especies propias del MSP que alcanzan el MSS por migración vertical (Juberthie 2000; Ledesma
et al. 2020; Giachino y Vailati 2010).
Hoy en día se reconocen diversos tipos de hábitat subterráneos poco profundos,
los denominados en inglés shallow subterranean habitats (SSH), entre los
cuales se encuentra el MSS (Culver y Pipan 2014).
El resultado final es un gradiente de vida que se expresa desde la superficie
del suelo, a través de los distintos horizontes edáficos, el MSS y otros SSH
hasta, finalmente, llegar al MSP (Gers 1998).
Aunque el MSS ha sido estudiado en diversas partes del
mundo, el continente europeo es, sin duda, la región geográfica que concentra
mayor número de trabajos (ver revisión en Mammola et
al. 2016). El objetivo del presente trabajo es, tras realizar una breve
descripción de este peculiar medio, ofrecer una revisión de los estudios que se
han llevado a cabo en la península ibérica y de los principales resultados
obtenidos.
Tipologías y características del MSS
El medio subterráneo superficial o MSS es, en esencia, el
conjunto de oquedades, grietas y fisuras que se encuentran en los niveles
profundos del suelo, normalmente en contacto con la roca madre. Al igual que
las cuevas, el MSS es un hábitat afótico en el que la amortiguación de las fluctuaciones
de temperatura y humedad con respecto a la superficie y, en general, los altos
niveles de humedad relativa (por ejemplo, Pipan et al.
2010; Nitzu et al. 2014; Rendoš et al. 2016; Barranco
et al. 2013; entre otros) propician la colonización de estos espacios
subterráneos por parte de fauna estenoica con un marcado carácter higrófilo.
Aunque se conocen aspectos básicos sobre la estructura y dinámica del MSS (ver
revisión en Mammola et al. 2016), todavía existe
poca información sobre la fauna que habita estos espacios subterráneos.
El MSS como fenómeno litológico-edafológico no es una rareza
paisajística y tampoco se restringe a un determinado paisaje o sustrato rocoso.
Mientras que se conocen MSS en paisajes kársticos que están íntimamente
relacionados con las grandes cavernas, las relaciones de otros MSS con las
grutas no parecen tan evidentes. Esta observación hace aún más interesante el
estudio de estos medios subterráneos ya que posibilitan la existencia de fauna
hipogea, incluso en paisajes silíceos que, en la mayoría de las ocasiones,
carecen de cuevas (Juberthie et al. 1980; Juberthie 2000).
Se han descrito cuatro tipos básicos de MSS según su origen.
En primer lugar, citamos el denominado MSS de roca madre, producto de la
meteorización química de la roca madre (Gers 1992),
siendo este un proceso paulatino y casi simultáneo a la formación de los
horizontes edáficos. Este tipo de MSS es típico de fondos de valle y, en
general, de superficies con poca o nula pendiente. Es, quizá, la tipología de
MSS menos explorada. En segundo lugar, está el MSS coluvial (Figs.
1 A-C y 2 A-D), también conocido
como MSS de ladera, característico de superficies montañosas con pendiente. Se
forma al depositarse fragmentos de roca cuyo origen está en la erosión mecánica
de los afloramientos rocosos (Juberthie et al.
1980). La naturaleza de la roca puede ser calcárea, silícea o volcánica, en
función de la naturaleza litológica de los afloramientos. Estos depósitos
rocosos pueden quedar cubiertos por un suelo más o menos potente, y sobre los
cuales puede medrar una cubierta vegetal en muy diversas etapas de evolución.
En otros casos el nivel superior del MSS queda desnudo, expuesto, mostrándose a
la vista como un canchal de montaña. Este tipo de MSS es uno de los más
estudiados desde el punto de vista faunístico (ver, por ejemplo: Růžička 1989; Rendoš et al.
2012; Ortuño et al. 2023; entre otros
muchos). El tercer tipo de MSS es el volcánico, resultado de la acumulación de
escorias volcánicas y piroclastos (Oromí et al. 1986; Oromí 2010; Macías-Hernández
et al. 2024) y cuya fauna ha sido ampliamente estudiada, por ejemplo, en
las Islas Canarias (ver Oromí et al. 2021; Macías-Hernández et al. 2024, entre otros).
Finalmente, el MSS más recientemente tipificado, el MSS aluvial (Figs. 1 D-E y 2 G)
(Ortuño et al. 2013), constituido por la red de
espacios subterráneos que se encuentran en los depósitos aluviales de cursos
fluviales temporales. Dado el carácter estacional de sus aguas, el lecho de los
cauces, y las terrazas aledañas, puede estar cubierto por un suelo más o menos
evolucionado, sobre el que pueden llegar a crecer plantas, tanto herbáceas como
leñosas. Estos depósitos se generan a partir de cualquier tipo de litología,
asentándose sobre la roca madre, y desarrollando una potencia que puede
alcanzar varios metros de espesor. Son MSS muy dinámicos que acogen fauna
terrestre y en los que, coincidiendo con los intervalos de actividad
hidrológica, también dan cabida a especies acuáticas (Ortuño
et al. 2013).

Figura 1. Algunos
enclaves ibéricos con depósitos coluviales y aluviales generadores de MSS: A)
coluvión calizo en La Serrella (Sistema Prebético alicantino); B)
coluvión silíceo en la Peña del Oso (Sistema Central); C) coluvión por
erosión, y deposición, de conglomerados en el Puerto de San Glorio (Montes
Cantábricos); D) MSS calizo sobre el que se ha formado suelo en terraza
fluvial en Vall d’Ebo (Sistema Prebético alicantino); E) aluvión calizo
en el río Ebo (Sistema Prebético alicantino).
Figure 1. Locations in the Iberian Peninsula with Some Iberian colluvial and
alluvial deposits that generate MSS: A) limestone colluvium in La
Serrella (Alicante, Prebetic System); B) siliceous colluvium in Peña del
Oso (Central System); C) colluvium due to erosion and deposition of
conglomerates in Puerto de San Glorio (Cantabrian Mountains); D)
limestone MSS on which a floodplain soil has formed in Vall d’Ebo (Alicante,
Prebetic System); E) limestone alluvium in the Ebo River (Alicante,
Prebetic System).
En el momento de valorar diferentes factores que contribuyen
a la diversidad faunística del MSS, y más allá de tener en cuenta la naturaleza
de la roca y las causas que lo formaron, es importante considerar la
posibilidad de que esa red de espacios subterráneos esté en contacto con las
mesocavernas y microcavernas (sensu Howart 1983)
del MSP. También es necesario considerar el papel que desempeña el desarrollo
de ciertos horizontes edáficos (horizontes A y B) sobre el MSS, o en las
proximidades de este, así como la presencia o ausencia de cubierta vegetal.
Otros factores como la localización geográfica, la altitud, la orientación, la
pendiente, la pluviosidad o la innivación (Fig. 2 E, F)
van a condicionar, junto a los ya mencionados, tanto los ciclos de temperatura
y de humedad en el MSS como la cantidad de materia orgánica que llega por
percolación desde el exterior, determinando finalmente las comunidades
faunísticas que van a ocupar esté hábitat subterráneo (Ortuño
et al. 2013; Ledesma et al. 2020; Eusebio et al. 2021; entre otros).

Figura 2. A, B) Instalación de una Estación de
Muestreo Subterráneo en MSS del Puerto de San Glorio (Montes Cantábricos); C)
sondeo en MSS del Puerto de Piedrasluengas (Montes Cantábricos); D)
instalación de una Estación de Muestreo Subterráneo en MSS del Puerto de la
Fuenfría (Sistema Central); E) accediendo en invierno a una Estación de
Muestreo Subterránea balizada en Dos Hermanas (Sistema Central); F)
accediendo en invierno a una Estación de Muestreo Subterránea no balizada en el
Alto de Navafría (Sistema Central); G) instalación de una Estación de
Muestreo Subterráneo en MSS aluvial en Tárbena (Sistema Prebético alicantino).
Figure 2. A, B) Installing a Subterranean Sampling Device in an MSS of Puerto de
San Glorio (Cantabrian Mountains); C) drilling in an MSS of Puerto de
Piedrasluengas (Cantabrian Mountains); D) placing a Subterranean
Sampling Device in an MSS of Puerto de la Fuenfría (Central System); E)
accessing in winter to a marked Subterranean Sampling Device in Dos Hermanas
(Central System); F) accessing in winter to an unmarked Subterranean
Sampling Device in Alto de Navafría (Central System); G) installing a
Subterranean Sampling Device in an alluvial MSS in Tárbena (Alicante, Prebetic
System).
El MSS en la península ibérica
En lo que a la península ibérica se refiere, la última
década ha sido testigo de un notable incremento de estudios detallados y
sistemáticos del MSS y su fauna (Fig. 3).
Entre las unidades de relieve de naturaleza calcárea
destacamos, en España, las prospecciones realizadas en las cordilleras
Prebética, Subbética y Penibética. En la Prebética se estudió, en el norte de
la provincia de Alicante, barrancos con MSS aluvial (Ortuño
et al. 2013), los cuáles interconectan montañas (El Montgó, Serra de
Bernia, Serra Aitana, La Serrella, Serra Mariola, Serra del Menejador, Penyes
Roset, Serra del Maigmó) en las que también se han estudiado ciertos aspectos
faunísticos y ecológicos del MSS coluvial (Gilgado
et al. 2015a, 2015b; Jiménez-Valverde et al. 2015; Akkari et al. 2018). En la Subbética, la atención
recayó en Sierra de María (Cifuentes y
Barranco 2020; Ortuño et al. 2020; Cifuentes et al. 2021), mientras que en la
Penibética se centró en la Sierra de Gádor (Cifuentes
et al. 2021). En la zona centro se muestreó el MSS en Tamajón (Ortuño et al. 2017). El MSS en el Prepirineo y los
Montes Cantábricos también está siendo prospectado (V.M. Ortuño, datos sin
publicar). En Portugal, el sistema kárstico de Arrábida ha sido objeto de
estudio con el fin de implementar técnicas cartográficas específicas para el
MSS (Eusebio et al. 2023). Otros MSS coluviales
localizados en las estribaciones occidentales del Sistema Central, en la
llamada Orla Occidental, también han sido explorados desde el punto de vista
faunístico (Eusebio et al. 2021), más
concretamente en el sistema montañoso Aire/Montejunto.
Con respecto a los relieves de
naturaleza silícea, hasta el momento los muestreos se han realizado en el
Sistema Ibérico, Sistema Central y la Cordillera Penibética, obteniéndose
resultados muy interesantes, algunos de los cuales han sido ya publicados. En
el Sistema Ibérico se está estudiando el MSS de dos enclaves, El Moncayo (Ortuño et al. 2014a; Gilgado
et al. 2014) y la Sierra de Mencilla. El MSS estudiado en el Sistema
Central está circunscrito a la Sierra de Guadarrama (Gilgado
et al. 2017; Baquero et al. 2017, 2021a, 2021b; Carles-Tolrá et al. 2018, 2019; Ledesma et al.
2019, 2020, 2022;
Ortuño et al. 2019, 2023;
Camacho y Ortuño 2019; Jordana et al. 2020) y Sierra de Ayllón (Ortuño y Toribio 1994; Barranco et al. 2013; Ortuño
et al. 2014b, 2023; Gilgado et al. 2015b). En la Cordillera Penibética los MSS silíceos que se han muestreado son
los de Sierra Nevada, concretamente en el Corral del Veleta (Gilgado et al. 2015b, 2015c;
Carles-Tolrá et al. 2018).
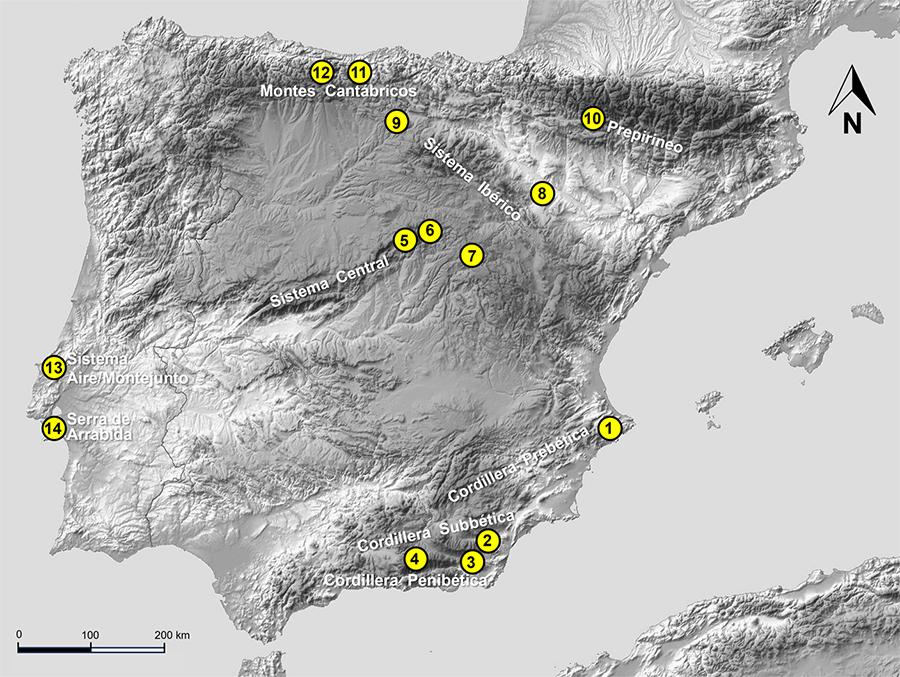
Figura 3. Áreas de la península ibérica en las
que se ha muestreado de forma sistemática el Medio Subterráneo Superficial
(MSS): 1) barrancos y sierras septentrionales de Alicante; 2)
Sierra de María (Almería); 3) Sierra de Gádor (Almería); 4)
Sierra Nevada (Granada); 5) Parque Nacional de la Sierra de Guadarrama
(Madrid/Segovia); 6) Sierra de Ayllón (Madrid); 7) Tamajón
(Guadalajara); 8) Sierra de El Moncayo (Zaragoza); 9) Sierra de
Mencilla (Burgos); 10) Sierra de Santo Domingo (Zaragoza); 11) La
Liébana (Cantabria); 12) Parque natural de Las Ubiñas-La Mesa
(Asturias); 13) sistema montañoso Aire/Montejunto (Leiría); 14)
sistema kárstico de Arrábida (Setúbal).
Figure 3. Locations of the Iberian Peninsula
where the MSS has been systematically sampled include: 1) canyons and
northern mountains of Alicante; 2) Sierra de María (Almería); 3)
Sierra de Gádor (Almería); 4) Sierra Nevada (Granada); 5) Sierra
de Guadarrama National Park (Madrid/Segovia); 6) Sierra de Ayllón
(Madrid); 7) Tamajón (Guadalajara); 8) Sierra de El Moncayo
(Zaragoza); 9) Sierra de Mencilla (Burgos); 10) Sierra de Santo
Domingo (Zaragoza); 11) La Liébana (Cantabria); 12) Las Ubiñas-La
Mesa Natural Park (Asturias); 13) Aire/Montejunto mountain system
(Leiría); 14) Arrábida karst system (Setúbal).
Novedades faunísticas para la península ibérica
El estudio sistematizado del MSS ha propiciado el
descubrimiento de especies e incluso géneros nuevos para la Ciencia, sin
olvidar que este tipo de prospecciones subterráneas también han brindado
registros de especies ya conocidas que, si bien sólo habían sido reportadas en
hábitats epigeos, ahora sabemos que también se hallan en el MSS y que incluso
este es su hábitat principal. En muchos casos, dada la elevada frecuencia de
aparición, hay que interpretarlas como especies troglófilas, o tendentes a la troglofilia.
En otros casos, especies que habían sido consideradas “cavernícolas” y con una
distribución geográfica muy puntual, se ha constatado que sus distribuciones no
están circunscritas a una o varias cuevas, sino que se hallan más extendidas, y
seguramente de forma continua, al haberse detectado en el MSS. A la luz de
todas estas novedades aparecidas, como resultado de los estudios sistemáticos e
intensivos realizados durante la última década, se ha visto incrementado de
manera significativa el conocimiento sobre la biodiversidad subterránea
ibérica, así como la autoecología y distribución de muchas especies. A
continuación, haremos un repaso a los descubrimientos faunísticos más notables.
Nuevos taxones (Fig. 4)
En lo que concierne al subfilo Hexapoda, el estudio del MSS en la Sierra de Guadarrama ha
arrojado 14 nuevas especies de Collembola (Baquero
et al. 2017, 2021a, 2021b; Jordana et al. 2020): Orchesella
mesovoides Baquero & Jordana, 2017; Orchesella colluvialis Jordana & Baquero, 2017; Friesea ortunoi Jordana & Baquero,
2020; Schaefferia sendrai Jordana & Baquero, 2020; Pachyotoma
penalarensis Baquero & Jordana, 2021; Entomobrya guadarramensis Jordana & Baquero, 2021; Entomobrya ledesmai Jordana & Baquero,
2021; Lepidocyrtus labyrinthi Baquero & Jordana, 2021; Lepidocyrtus
paralignorum Baquero & Jordana, 2021; Lepidocyrtus purgatori Baquero & Jordana, 2021; Pseudosinella valverdei Baquero & Jordana,
2021; Pseudosinella gonzaloi Baquero & Jordana, 2021; Pygmarrhopalites
custodum Baquero & Jordana, 2021; y Allacma cryptica Baquero
& Jordana, 2021. Por otra parte, se ha descrito una nueva especie de
Diplura en la Cordillera Prebética alicantina, Campodea (Campodea)
alluvialis Sendra, 2017 (Sendra et al.
2017), y dos especies de Insecta (Orthoptera y
Coleoptera) en la Sierra de Ayllón y El Moncayo, el grillo Nemobius
interstitialis Barranco, Gilgado & Ortuño, 2013 (Barranco et al. 2013) y
el carábido Trechus arrecheai Ortuño, Gilgado & Cuesta, 2014 (Ortuño et al. 2014a).

Figura 4. Especies descritas del Medio
Subterráneo Superficial (MSS): A, B) Espécimen completo y detalle de la
cabeza de Trechus arrecheai Ortuño, Gilgado & Cuesta, 2014
(endemismo del El Moncayo); C) Nemobius interstitialis Barranco,
Gilgado & Ortuño, 2013 (endemismo del este del Sistema Central); D) Guadarramasoma
ramosae Gilgado, Ledesma, Enghoff & Mauriès, 2017 (endemismo de la
Sierra de Guadarrama); E) Archipolydesmus altibaeticus Gilgado &
Enghoff, 2015 (endemismo de Sierra Nevada). Barra de escalas: A) 1 mm; B)
0,5 mm; C) 5 mm; D-E) 2 mm.
Figure 4. Some new species discovered in the
MSS: A, B) Trechus arrecheai Ortuño, Gilgado & Cuesta, 2014
(endemic to El Moncayo); C) Nemobius interstitialis Barranco,
Gilgado & Ortuño, 2013 (endemic to the eastern Central System); D) Guadarramasoma
ramosae Gilgado, Ledesma, Enghoff & Mauriès, 2017 (endemic to Sierra de
Guadarrama); E) Archipolydesmus altibaeticus Gilgado &
Enghoff, 2015 (endemic to Sierra Nevada). Scale bars: A)
1 mm; B) 0.5 mm; C) 5 mm; D-E) 2 mm.
Ampliación sobre el conocimiento de especies ya
conocidas (Figs. 5
y 6)
El subfilo Crustacea ha aportado,
por el momento, una novedad procedente de la Sierra de María, Porcellio
selomai Cifuentes & Barranco, 2020 (Cifuentes y Barranco 2020).
Respecto al subfilo Myriapoda, son 5 las especies nuevas de
diplópodos que se han descrito procedentes de las Prebéticas alicantinas,
Sierra Nevada, Sierra de Ayllón y Sierra de Guadarrama (Gilgado et al. 2015b, 2015c;
2017; Akari et al.
2018): Archipolydesmus foliatus Gilgado & Enghoff, 2015; Archipolydesmus
altibaeticus Gilgado & Enghoff, 2015; Ceratosphys cryodeserti
Gilgado, Mauriès & Enghoff, 2015; Guadarramasoma ramosae Gilgado,
Ledesma, Enghoff & Mauriès, 2017 (siendo también nuevo género); y
Ommatoiulus longicornis Akkari & Enghoff, 2018.
Dentro del subfilo Hexapoda se han hallado dos especies de
Diplura, Campodea propinqua Silvestri, 1932 y Campodea neusae
Sendra & Moreno, 2006, por primera vez en la Sierra de Guadarrama, y
también en el MSS, modificando con diferente amplitud su distribución
inicialmente conocida (Sendra et al. 2017): C.
propinqua antes sólo conocida en el Sistema Bético; C. neusae
inicialmente conocida en la Sierra de Ayllón. Las prospecciones en el MSS han
permitido colectar numerosos Zygentoma del complejo específico de Coletinia
maggii (Grassi, 1887), proporcionado nuevas localidades, que han de
facilitar un mejor conocimiento de la composición taxonómica de este grupo de
especies, así como de los rangos de variabilidad (Gilgado
y Ortuño 2015).
Entre los Coleoptera, son
muy diversos los ejemplos que se manejan de cambios en el conocimiento
corológico y autoecológico de las especies. Por ejemplo: el Elateridae Dima assoi Pérez Arcas, 1872, cuya presencia regular de los imagos en el subsuelo
de El Moncayo permite ampliar aspectos sobre su biología (Gilgado et al. 2014), o la constatación de la presencia en un MSS
aluvial del Carabidae, Trechus fulvus Dejean, 1831, lo que permite
aventurar la hipótesis de que este medio pueda ser utilizado por esta especie,
y por otras, como corredor subterráneo, facilitado así la dispersión de
artrópodos higrófilos que, a priori, cuentan con bajo potencial de dispersión
en el medio epigeo (Ortuño et al. 2017). Cuatro
Carabidae epigeos fueron observados como integrantes regulares de las
comunidades hipogeas del MSS de la Sierra de Guadarrama, Leistus (Leistus)
constrictus Schaufuss, 1862; Nebria (Nebria) vuillefroyi Chaudoir,
1866; Trechus (Trechus) schaufussi pandellei Putzeys, 1870; y Laemostenus
(Eucryptotrichus) pinicola (Graells, 1851); lo que condujo a redefinir su
autoecología, y a ponderar el protagonismo hipogeo de las “larvas vs
imagos” de cada una de estas especies (Ortuño et al. 2019). Con un foco geográfico más amplio, se evaluó la
distribución del Carabidae epigeo Leistus (Pogonophorus) puncticeps
Fairmaire & Laboulbène, 1854, observándose la presencia de imagos en el
MSS, tanto en enclaves de clima eurosiberiano (El Moncayo), coincidente con el
área de distribución previamente conocida, como en nuevos enclaves meridionales
de clima mediterráneo (formaciones montañosas del norte de la provincia de
Alicante y Sierra de María). Esto pone de manifiesto la tendencia subtroglófila
de esta especie y subraya la relevancia del MSS como refugio de fauna higrófila
en regiones meridionales más xéricas (Ortuño et al. 2020). Otra familia de Coleoptera con importante
representación en el MSS son los Leiodidae, los cuáles se estudiaron de un modo
exhaustivo en su contribución a las comunidades hipogeas de la Sierra de
Guadarrama. Del conjunto de especie destacamos cuatro, por ser el primer
registro de ellas en la Sierra de Guadarrama, y para algunas también el primer
dato sobre su presencia en el MSS (Ledesma et al. 2022): Attumbra josephinae josephinae (Saulcy,
1862); Catops fuliginosus Erichson, 1837; Catopsimorphus
(Weiratherella) rougeti Saulcy, 1864; y Choleva (Choleva) cisteloides
cisteloides (Frölich, 1799), esta última también novedosa para el Sistema
Central.
Un interesante Hymenoptera de la familia Formicidae, Aphaenogaster cardenai Espadaler, 1981, fue
hallado en el MSS de la Sierra de Ayllón, lo que ampliaba su registro desde la
mitad meridional de la península ibérica hasta el Sistema Central, y aportaba
el primer registro subterráneo de esta hormiga, no en cuevas, ubicándola en un
entorno orográfico de carácter silíceo (Ortuño et
al. 2014b). Los Diptera también se han significado como un orden de Insecta
que están bien representados en diversidad y abundancia en el subsuelo.
Destacamos cuatro especies que han resultado novedosas para la fauna española
y/o ibérica, así como en su vinculación a medios hipogeos, más concretamente al
MSS (Carles-Tolrá et al. 2018, 2019): en Sierra Nevada y Sierra de Guadarrama,
Crumomyia glacialis (Meigen, 1830); en Sierra de Guadarrama, Anapausis
helvetica Haenni, 1984, Crumomyia gelida (Hackman, 1965), y Crumomyia
notabilis (Collin, 1902).
Insectos hemimetábolos también tienen una interesante
presencia en el MSS y se empieza a disponer de ciertos datos que amplían el
conocimiento de algunas de sus especies. Tal es el caso del Orthoptera Paramogoplistes
novaki (Krauss, 1888), que fue hallado en el Sistema Prebético alicantino,
siendo esta la primera referencia de esta especie para el ámbito ibérico (Barranco y Ortuño 2021); su localización no
exclusiva en el MSS indica que esta especie además de tener hábitos epigeos
también circula por los intersticios del subsuelo.
Aunque el subfilo
Crustacea no ha proporcionado tanta información como los demás subfilos, la
presencia de dos especies de Bathynellacea, Hexaiberobathynella mateusi
(Galhano, 1967) y Hexabathynella nicoleiana Camacho, 1986, más allá de
las novedades respecto a su distribución (ambas especies son novedad en
elevaciones de la Sierra de Guadarrama) y autoecología, han propiciado que se
contemple al MSS como un medio que, en determinadas circunstancias, puede ser
muy sumamente cambiante. Su captura en MSS coluviales resalta la cualidad de
que algunos MSS puedan mutar de ser espacios subterráneos terrestres a,
temporalmente, ambientes subterráneos inundados por elevación del nivel
freático (Camacho y Ortuño 2019).
El subfilo Myriapoda,
hasta el momento, aportó novedades corológicas y autoecológicas de cuatro
especies de Diplopoda. Psichrosoma baeticaense (Mauriès, 2013) (antes Typhlopsychrosoma
baeticaense) del que se conocía su presencia en cuatro cuevas del sistema
Bético, fue hallado en el MSS del Sistema Prebético alicantino ampliando así su
área distribución, y dejando entrever que esta especie debe tener una
distribución continua a través del medio subterráneo (Gilgado et al. 2015a). Archipolydesmus bedeli (Brölemann, 1902),
sólo se conocía a partir de dos registros en la Sierra de Guadarrama con un
exiguo número de ejemplares, pero su colecta numerosa en el MSS de la Sierra de
Ayllón sirvió para ampliar su conocimiento corológico, autoecológico, y permitió
una detallada redescripción de la especie (Gilgado et al. 2015b). En el Sistema Penibético, concretamente en
Sierra Nevada, dos especies conocidas en la zona se registran por primera vez,
y de forma abundante, en el MSS, Proteroiulus hispanus Schubart, 1959 y Ommatoiulus
ilicis (Brölemann, 1897), quizá como medio alternativo en donde hallar la
humedad que no encuentran regularmente con hábitos de vida epigeos (Gilgado et al. 2015c).
Por último, el subfilo Chelicerata, ha proporcionado
interesantes hallazgos en muy diferentes enclaves ibéricos, y como ejemplo de
ello citamos Cybaeodes dosaguas Ribera y De Mas, 2015, araña sólo
conocida a partir de dos hembras colectadas en una cueva de la provincia de
Valencia (Ribera y De Mas 2015) y de la que
ahora tenemos constancia de su presencia en el MSS del Sistema Prebético
alicantino. En el MSS de la Sierra de Ayllón se constató la abundante presencia
de Harpactocrates gurdus Simon, 1914, tanto adultos como juveniles, lo
que indica que estos medios subterráneos también forman parte de sus hábitos de
vida. Un estudio pormenorizado de la fauna del MSS de la Sierra de Guadarrama,
ha proporcionado 32 especies de arañas novedosas para el Parque Nacional de la
Sierra de Guadarrama, de las cuáles dos, Liocranum apertum Denis, 1960 y
Walckenaeria capito (Westring, 1861), eran desconocidas para el ámbito
peninsular (Ledesma et al. 2019). Además, las
citas para casi el 50% de las especies halladas en el MSS del Parque Nacional
suponen un significativo aumento en su rango de distribución ibérica (Ledesma et al. 2019). Finalmente, los autores de
este estudio clasificaron al gnafósido Drassodex granja Hervé, Roberts
& Murphy, 2009 como especie troglófila (conocida hasta ahora de un solo
registro antiguo), pero relativamente común en el MSS de Guadarrama.
Los párrafos anteriores son
sólo una muestra necesariamente sucinta de un conjunto de información mucho más
amplia y profunda sobre la distribución y autoecología de las especies que el
lector interesado puede hallar en las publicaciones citadas.

Figura 5. Especies halladas en el Medio
Subterráneo Superficial (MSS): A) Dima assoi Pérez Arcas, 1872
(El Moncayo); B) Trechus (Trechus) fulvus Dejean, 1831 (Tamajón);
C) Nebria (Nebria) vuillefroyi Chaudoir, 1866 (Sierra de
Guadarrama); D) Catops fuliginosus Erichson, 1837 (Sierra de
Guadarrama); E) Aphaenogaster cardenai Espadaler, 1981 (Sierra de
Ayllón); F) Hexaiberobathynella mateusi (Galhano, 1967) (Sierra
de Guadarrama). Barra de escalas: A, B, E) 2 mm;
C) 3 mm; D) 1 mm; F) 0,25 mm.
Figure 5. Some species found in the MSS: A) Dima assoi Pérez Arcas, 1872 (El Moncayo); B) Trechus
(Trechus) fulvus Dejean, 1831 (Tamajón); C) Nebria (Nebria) vuillefroyi Chaudoir, 1866 (Sierra de Guadarrama); D) Catops fuliginosus Erichson, 1837 (Sierra de Guadarrama); E) Aphaenogaster cardenai Espadaler, 1981 (Sierra de Ayllón); F) Hexaiberobathynella mateusi (Galhano, 1967) (Sierra de Guadarrama). Scale bars: A, B, E)
2 mm; C) 3 mm; D) 1 mm; F) 0.25 mm.

Figura 6. Especies halladas en el Medio
Subterráneo Superficial (MSS): A) Psichrosoma baeticaense
(Mauriès, 2013) (Sistema Prebético); B) Archipolydesmus bedeli
(Brölemann, 1902) (Sierra de Ayllón); C) Cybaeodes dosaguas
Ribera & De Mas, 2015 (Sistema Prebético); D-F) macho, hembra y
juvenil de Harpactocrates gurdus Simon, 1914 (Sierra de Ayllón). Barra
de escalas: A-F) 2 mm.
Figure 6. Some species found in the MSS: A)
Psichrosoma baeticaense (Mauriès, 2013) (Sistema Prebético); B) Archipolydesmus
bedeli (Brölemann, 1902) (Sierra de Ayllón); C) Cybaeodes dosaguas
Ribera & De Mas, 2015 (Sistema Prebético); D-F) male, female and
juvenile of Harpactocrates gurdus Simon, 1914 (Sierra de Ayllón). Scale
bars: A-F) 2 mm.
Conclusiones
Los déficits en el conocimiento de la biodiversidad suponen
el primer impedimento para su correcta protección (ver una revisión en Hortal et al. 2015). El déficit más importante es el
denominado déficit Linneano, el cual hace referencia a la diferencia que existe
entre el número real de especies existentes y el número de especies actualmente
descritas (Lomolino 2004). Es el déficit más
importante ya que condiciona a todos los demás (Hortal
et al. 2015). El segundo déficit más relevante es el déficit Wallaceano, el
cual hace referencia a la falta de conocimiento que tenemos sobre la
distribución geográfica de las especies (Lomolino 2004).
El tercer déficit que queremos señalar aquí es el denominado déficit
Hutchinsoniano, el cual hace referencia a la falta de conocimiento acerca de
los requerimientos ambientales de las especies (Cardoso
et al. 2011). Como queda patente en el presente
trabajo, el estudio sistematizado e intensivo del medio subterráneo en general,
y del MSS en particular, puede proporcionar avances significativos en la
reducción de estos déficits del conocimiento. Sin embargo, el medio
subterráneo, debido a su condición de medio inhóspito, oculto y desconocido, se
enfrenta a no pocas amenazas y es urgente adoptar medidas para su protección (Mammola et al. 2019).
Contribución de los autores
Ambos
autores (V.M.O. y A.J.V.) han contribuido por igual en la concepción del
artículo y en su redacción. Las figuras son obra de V.M.O.
Agradecimientos
Queremos expresar nuestro agradecimiento al Dr. José D.
Gilgado por la cesión de alguna de las imágenes que ilustran este trabajo.
Referencias
Akkari, N., Gilgado, J.D.,
Ortuño, V.M., Enghoff, H. 2018. Out of the dark void: Ommatoiulus
longicornis n. sp., a new julid from Spain (Diplopoda, Julida) with notes
on some troglobiomorphic traits in millipedes. Zootaxa 4420(3):
415-429. https://doi.org/10.11646/zootaxa.4420.3.7
Baquero, E., Ledesma, E.,
Gilgado, J.D., Ortuño, V.M., Jordana, R. 2017. Distinctive
Collembola communities in the Mesovoid Shallow Substratum: First data for the
Sierra de Guadarrama National Park (Central Spain) and a description of two new
species of Orchesella (Entomobryidae). PLoS ONE 12(12):
e0189205. https://doi.org/10.1371/journal.pone.0189205
Baquero, E., Jordana,
R., Ortuño, V.M. 2021a. Distinctive
Collembola Communities in the Mesovoid Shallow Substratum: Entomobryomorpha of the Sierra de Guadarrama National Park (Central Spain). Zoosystema
43(3): 37-78. https://doi.org/10.5252/zoosystema2021v43a3
Baquero, E., Jordana,
R., Ortuño, V.M. 2021b. Neelipleona and Symphypleona
(Collembola) from a sampling in the mesovoid shallow substratum of the Sierra
de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and
Biogeography. Insects 12: 266. https://doi.org/10.3390/insects12030266
Barranco, P., Ortuño,
V.M. 2021. Nuevos datos de Mogoplistidae (Insecta, Orthoptera) y primer
registro de Paramogoplistes novaki (Krauss, 1888) en la península
ibérica. Graellsia 77(2): e148. https://doi.org/10.3989/graellsia.2021.v77.309
Barranco, P., Gilgado,
J.D., Ortuño, V.M. 2013. A new mute species of the genus Nemobius
Serville (Orthoptera, Gryllidae, Nemobiinae) discovered in colluvial, stony
debris in the Iberian Peninsula: A biological, phenological and biometric
study. Zootaxa 3691(2): 201-219. https://doi.org/10.11646/zootaxa.3691.2.1
Bonet, F. 1931. Estudios sobre
Collembolos cavernícolas con especial referencia a los de la española. Memorias
de la Sociedad española de Historia Natural. Madrid 14(4): 231-403.
Camacho,
A.I., Ortuño, V.M. 2019. Unusual habitat for Bathynellacea
(Crustacea, Malacostraca): first record of this groundwater crustacean in the
mesovoid shallow substratum (MSS). Graellsia 75(2): e100. https://doi.org/10.3989/graellsia.2019.v75.248
Cardoso, P., Erwin, T.L., Borges, P.A., New, T.R. 2011. The seven
impediments in invertebrate conservation and how to overcome them. Biological
Conservation 144: 2647-55. https://doi.org/10.1016/j.biocon.2011.07.024
Carles-Tolrá,
M., Tinaut, A., Gilgado, J.D., Ortuño, V.M. 2018. Presence of Crumomyia glacialis (Meigen, 1830) (Diptera:
Sphaeroceridae) in the Sierra Nevada and Sierra de Guadarrama National Parks
(Spain): first record from the Iberian Peninsula, and evidence of its presence
in the Mesovoid Shallow Substratum. Boletín de la Sociedad
Entomológica Aragonesa 63: 182-186.
Carles-Tolrá, M.,
Jiménez-Valverde, A., Ledesma, E., Ortuño, V.M. 2019. Three
new dipteran species newly recorded in the Mesovoid Shallow Substratum in the
Sierra de Guadarrama National Park, Spain (Diptera: Scatopsidae and
Sphaeroceridae). Boletín de la Sociedad Entomológica Aragonesa
64: 137-142.
Casale, A., Rondolini,
G. 1983. Morphologie, distribution, écologie d'une espèce de
Bathysciinae en Milieu Souterrain Superficiel au Piémont (Coleoptera,
Catopidae). Mémoires de Biospéologie 10: 73-82.
Cifuentes, J., Barranco, P. 2020. Porcellio
selomai sp. n. (Oniscidea, Porcellionidae), un nuevo isópodo terrestre del
medio subterráneo superficial (MSS) de la península ibérica. Boletín de la
Asociación Española de Entomología 44(1-2): 167-182.
Cifuentes, J., López
Martínez, S., Molina Pardo, J.L., Barranco, P. 2021. Isópodos (Crustacea,
Oniscidea) del Medio Subterráneo Superficial (MSS) en el Sureste Ibérico
(Almería, España). Boletín de la Asociación Española de Entomología
45(3-4): 261-276.
Culver, D.C., Pipan, T.
2014. Shallow Subterranean Habitats: Ecology, Evolution,
and Conservation. Oxford University Press. Oxford,
UK. https://doi.org/10.1093/acprof:oso/9780199646173.001.0001
Delić, T., Trontelj,
P., Zakśek, V., Fišer, C. 2016. Biotic and abiotic determinants of appendage
length evolution in a cave amphipod. Journal of Zoology 299: 42-50. https://doi.org/10.1111/jzo.12318
Eusébio, R.P., Enghoff, H., Solodovnikov, A., Michelsen, A., Barranco, P.,
Salgado, J.M., Sendra, A., et al. 2021. Temporal and
spatial dynamics of arthropod groups in terrestrial subsurface habitats in
central Portugal. Zoology 147: 125931. https://doi.org/10.1016/j.zool.2021.125931
Eusébio, R.P., Fonseca,
P.E., Rebelo, R., Da Luz Mathias, M., Reboleira, A.S.P.S. 2023. How to map potential mesovoid shallow substratum (MSS) habitats? A
case study in colluvial MSS. Subterranean Biology 45: 141-156. https://doi.org/10.3897/subtbiol.45.96332
Fresneda, J., Hernando, C.,
Lagar, A., Ortuño, V.M. 1997. Sistemática y geonemia de un coleóptero
subterráneo de España: Oscadytes rovirai Lagar, 1975 (Coleoptera:
Pterostichidae). Annales de la Société Entomologique de France
(Nouvelle Série) 33(2): 205-213. https://doi.org/10.1080/21686351.1997.12277752
Gers, C. 1992. Ecologie et biologie des populations d’Arthropodes
terrestres du milieu Souterrain Superficiel: fonctionnement et écologie
évolutive. Tesis. Universidad Paul Sabatier,
Toulouse, France.
Gers, C. 1998. Diversity of energy fluxes and interactions between
arthropod communities: from Soil to Cave. Acta
Oecologica 19(3): 205-213. https://doi.org/10.1016/S1146-609X(98)80025-8
Giachino,
P.M., Vailati, D. 2010. The subterranean
environment. Hypogean life, concepts and collecting techniques. WBA
Handbooks, Verona, Italy.
Gilgado, J.D.,
Ortuño, V.M. 2015. Intra- and
inter-population polymorphism in Coletinia maggii (Grassi, 1887)
(Zygentoma: Nicoletiidae), an inhabitant of soil, mesovoid shallow substratum
(MSS) and caves—A challenge for the strict classification of subterranean
fauna? Zootaxa 3920(1): 085-100. https://doi.org/10.11646/zootaxa.3920.1.5
Gilgado, J.D., Ledesma, E.,
Cuesta, E., Arrechea, E., Zapata, J.L., Sánchez-Ruiz, A., Ortuño, V.M. 2014. Dima assoi Pérez Arcas 1872 (Coleoptera:
Elateridae): from montane to hypogean life. An example of exaptations to the
subterranean environment? Annales de la Société
entomologique de France (N.S.) 50(3-4): 264-271. https://doi.org/10.1080/00379271.2014.981421
Gilgado, J.D., Enghoff, H.,
Ortuño, V.M. 2015a. The hypogean Iberian genus Typhlopsychrosoma
Mauriès, 1982 (Diplopoda, Chordeumatida, Vandeleumatidae): distribution map,
key to species, first record in a Mesovoid Shallow Substratum (MSS) and
detailed iconography of T. baeticaense (Mauriès, 2013). Zootaxa
3937(2): 337-346. https://doi.org/10.11646/zootaxa.3937.2.5
Gilgado, J.D., Enghoff, H., Tinaut, A., Ortuño, V.M. 2015b. Hidden biodiversity in the Iberian Mesovoid Shallow Substratum
(MSS): New and poorly known species of the millipede genus Archipolydesmus
Attems, 1898 (Diplopoda, Polydesmidae). Zoologischer Anzeiger 258:
13-38. https://doi.org/10.1016/j.jcz.2015.06.001
Gilgado, J.D., Enghoff, H., Tinaut, A., Mauriès, J.–P., Ortuño, V.M. 2015c.
Sierra Nevada (Granada, Spain): a high-altitude biogeographical crossroads for
millipedes (Diplopoda), with first data on its MSS fauna and description of a
new species of the genus Ceratosphys Ribaut, 1920 (Chordeumatida:
Opisthocheiridae). Zootaxa 4044(3): 391-410. https://doi.org/10.11646/zootaxa.4044.3.4
Gilgado, J.D., Ledesma, E., Enghoff, H., Mauriès, J.-P., Ortuño, V.M. 2017. A
new genus and species of Haplobainosomatidae (Diplopoda: Chordeumatida) from
the MSS of the Sierra de Guadarrama National Park, central Spain. Zootaxa
4347(3): 492-510. https://doi.org/10.11646/zootaxa.4347.3.4
Hortal, J., de Bello, F., Diniz-Filho, J.A.F., Lewinsohn, T.M., Lobo, J.M.,
Ladle, R.J. 2015. Seven shortfalls that beset
large-scale knowledge of biodiversity. Annual Review of Ecology, Evolution,
and Systematics 46: 523-549. https://doi.org/10.1146/annurev-ecolsys-112414-054400
Howarth, F.G. 1983. Ecology of cave Arthropods. Annual Review of.
Entomology 28: 365-389. https://doi.org/10.1146/annurev.en.28.010183.002053
Jeannel, R. 1926. Faune cavernicole de la France, avec une étude des
conditions d’existence dans le domaine souterrain. Encyclopédie
Entomologique, 7. Lechevalier, Paris, France.
Jeannel, R. 1943. Les fossiles vivants des cavernes. L'avenir de la
science (n.s.), 1, Gallimard. Paris, France.
Jiménez-Valverde, A., Gilgado, J.D., Sendra, A., Pérez-Suárez, G., Herrero-Borgoñón,
J.J., Ortuño, V.M. 2015. Exceptional invertebrate
diversity in a scree slope in Eastern Spain. Journal of Insect Conservation
19: 713-728. https://doi.org/10.1007/s10841-015-9794-1
Jordana, R., Baquero, E., Ledesma, E., Sendra, A., Ortuño, V.M. 2020. Poduromorpha
(Collembola) from a sampling in the mesovoid shallow substratum of the Sierra
de Guadarrama National Park (Madrid and Segovia, Spain): Taxonomy and
Biogeography. Zoologischer Anzeiger 285: 81-96. https://doi.org/10.1016/j.jcz.2020.02.001
Juberthie, Ch. 1983. Le milieu souterrain: étendue et composition. Mémoires
de Biospéologie 10: 17-65.
Juberthie, Ch. 2000. The diversity of the karstic and pseudokarstic hypogean
habitats in the world. En: Wilkens, H., Culver, D.C., Humphreys, W. F. (eds.).
Ecosystems of the World 30. Subterranean Ecosystems, Pp. 17-39.
Elsevier, Amsterdan, The Netherlands.
Juberthie, Ch., Delay, D., Bouillon, M. 1980. Extension du milieu souterrain en
zone non calcaire: description d’un nouveau milieu et de son peuplement par les
Coléoptères troglobies. Mémoires de Biospéologie 7: 19-52.
Juberthie, Ch., Bouillon,
M., Delay, B. 1981. Sur l'existence du milieu souterrain
superficiel en zone calcaire. Mémoires de Biospéologie 8: 77-93.
Ledesma, E.,
Jiménez-Valverde, A., De Castro, A., Aguado-Aranda, P., Ortuño, V.M. 2019. The study of hidden habitats sheds light on poorly known taxa:
spiders of the Mesovoid Shallow Substratum. ZooKeys 841: 39-59. https://doi.org/10.3897/zookeys.841.33271
Ledesma, E.,
Jiménez-Valverde, A., Baquero, E., Jordana, R., De Castro, A., Ortuño, V.M.
2020. Arthropod biodiversity patterns point to the Mesovoid
Shallow Substratum (MSS) as a climate refugium. Zoology 141: 1-8.
https://doi.org/10.1016/j.zool.2020.125771
Ledesma, E.,
Jiménez-Valverde, A., Salgado, J.M., Ortuño, V.M. 2022. Cholevinae
(Coleoptera: Leiodidae) of the Sierra de Guadarrama National Park, Spain:
occurrence in the MSS of a siliceous landscape. Zoosystema 44(4):
125-149. https://doi.org/10.5252/zoosystema2022v44a4
Lomolino, M.V. 2004. Conservation biogeography. En: MV Lomolino, LR
Heaney (eds.). Frontiers of Biogeography: New Directions in
the Geography of Nature, pp. 293-96. Sinauer,
Sunderland, MA, USA.
Macias-Hernández,
N., Suárez, D., de la Cruz López, S., López, H., Oromí, P. 2024. Diversidad de
arañas hipogeas del archipiélago canario. Ecosistemas 2516. https://doi.org/10.7818/ECOS.2516
Mammola, S., Giachino, P.M., Piano, E., Jones, A., Barberis, M., Badino,
G., Isaia, M. 2016. Ecology and sampling techniques of an understudied
subterranean habitat: the Milieu Souterrain Superficiel (MSS). The
Science of Nature 103: 88. https://doi.org/10.1007/s00114-016-1413-9
Mammola, S., Cardoso, P.,
Culver, D. C., Deharveng, L., Ferreira, R. L., Fišer, C., et al. 2019. Scientists' warning on the conservation of subterranean ecosystems. BioScience
69: 641-650. https://doi.org/10.1093/biosci/biz064
Martel, E.A. 1908. L'Évolution Souterraine. Flammarion,
Paris, France.
Moseley, M. 2010. Are all caves ecotones? Cave and karst science 36(2009):
53-58.
Nitzu, E., Nae, A., Băncilă, R., Popa, I., Giurginca, A., Plăiaşu, R. 2014. Scree habitats: ecological function, species conservation and
spatial-temporal variation in the arthropod community. Systematics
and Biodiversity 12: 65-75. https://doi.org/10.1080/14772000.2013.878766
Oromí, P. 2010. La fauna
subterránea de Canarias: un viaje desde las lavas hasta las cuevas. En: Alfonso
Carrillo J. (ed.). Volcanes, mensajeros del fuego, creadores de vida,
forjadores del paisaje, pp. 63-98. Instituto de Estudios Hispánicos de
Canarias, Puerto de la Cruz, España.
Oromi, P., Medina, A.L.,
Tejedor, M.L. 1986. On the existence of
a superficial underground compartment in the Canary Islands. Acta del
IX Congreso Internacional de Espeleología. Barcelona 2: 147-151.
Oromí, P., Arechavaleta, M.A.,
De La Cruz, S., García, R., Izquierdo, I., López, H., et al. 2021. Diversidad
faunística en el medio subterráneo volcánico, con especial énfasis en las Islas
Canarias. Boletín de la Sociedad Española de Espeleología y Ciencias del
Karst (SEDECK) 16: 25-50.
Ortuño, V.M. 1996. Nuevos datos
sobre Caraboidea de la Península Ibérica (2ª nota). Boletín de la Asociación
española de Entomología 20(1-2): 193-200.
Ortuño, V.M. 2015. Los
Artrópodos en el contexto del bosque como refugio climático. Capítulo 11. pp:
171-184. En: Herrero, A., Zavala, M.A. (eds.), Los Bosques y la
Biodiversidad frente al Cambio Climático: Impactos, Vulnerabilidades y
adaptación en España. Ministerio de Agricultura, Alimentación y Medio
Ambiente. Madrid, España. 610 pp.
Ortuño, V.M., Toribio,
M. 1994. Nuevos datos sobre la distribución y ecología de Nebria vuillefroyi
Chaudoir, 1866 (Col., Nebriidae). Boletín de la Asociación española de
Entomología 18(1-2): 204-205.
Ortuño, V.M., Gilgado, J.D.,
Jiménez-Valverde, A., Sendra, A., Pérez-Suárez, G., Herrero-Borgoñón, J.J.
2013. The "Alluvial Mesovoid Shallow Substratum", a
New Subterranean Habitat. PLoS ONE 8(10): e76311. https://doi.org/10.1371/journal.pone.0076311
Ortuño, V.M., Cuesta, E.,
Gilgado, J.D., Ledesma, E. 2014a. A new hypogean Trechus
Clairville (Coleoptera, Carabidae, Trechini) discovered in a non-calcareous
Superficial Subterranean Habitat of the Iberian System (Central Spain). Zootaxa
3802(3): 359-372. https://doi.org/10.11646/zootaxa.3802.3.5
Ortuño, V.M., Gilgado, J.D.,
Tinaut, A. 2014b. Subterranean Ants: The Case of Aphaenogaster
cardenai (Hymenoptera: Formicidae). Journal of
Insect Science 14(212): 1-7. https://doi.org/10.1093/jisesa/ieu074
Ortuño, V.M., Ledesma, E., Gilgado, J.D., Veguillas, L., Barranco, P.
2017. On the distribution and autoecology of Trechus
fulvus Dejean, 1831 (Coleoptera: Carabidae: Trechinae) in the Iberian
Peninsula. Boletín de la Sociedad Entomológica Aragonesa 60:
195-206.
Ortuño, V.M., Ledesma, E.,
Jiménez-Valverde, A., Pérez-Suárez, G. 2019. Studies of the
mesovoid shallow substratum can change the accepted autecology of species: the
case of ground beetles (Coleoptera: Carabidae) in the Sierra de Guadarrama
National Park (Spain). Animal Biodiversity and Conservation
42(2): 213-226. https://doi.org/10.32800/abc.2019.42.0213
Ortuño, V.M., Barranco, P.,
Jiménez-Valverde, A., Sendra, A. 2020. El relicto glacial Leistus
(Pogonophorus) puncticeps Fairmaire & Laboulbène, 1854 (Coleoptera,
Carabidae): nuevos datos sobre distribución, autoecología y presencia en el
Medio Subterráneo Superficial (MSS). Graellsia 76(1): e107. https://doi.org/10.3989/graellsia.2020.v76.255
Ortuño, V.M.,
Jiménez-Valverde, A., Baquero, E., Jordana, R., Ledesma, E., Pérez-Suárez, G.,
Sendra, A., et al. 2023. Fauna del medio subterráneo superficial (MSS) en el
Parque Nacional de la Sierra de Guadarrama (España), pp: 49-76, En: Proyectos
de investigación en Parques Nacionales: 2015-2019. Organismo Autónomo de
parques Nacionales. Madrid, España. 407 pp.
Pipan, T. López, H., Oromí,
P., Polak, S., Culver, D.C. 2010. Temperature variation and
the presence of troglobionts in terrestrial shallow subterranean habitats. Journal
of Natural History 45: 253-273. https://doi.org/10.1080/00222933.2010.523797
Racovitza, E.G. 1907. Éssai sur les problèmes
biospéologiques. Archives de Zoologie Expérimentale et Générale. Serie 4 6: 371-488.
Rendoš, M., Mock, A., Jászay, T. 2012. Spatial and temporal dynamics of
invertebrates dwelling karstic mesovoid shallow substratum of Sivec National
Nature Reserve (Slovakia), with emphasis on Coleoptera. Biologia 67:
1143-1151. https://doi.org/10.2478/s11756-012-0113-y
Rendoš, M., Raschmanová, N., Kováč, Ľ., Miklisová, D., Mock, A., Ľuptáčik,
P. 2016. Organic carbon content and temperature as substantial factors
affecting diversity and vertical distribution of Collembola on forested scree
slopes. European Journal of Soil Biology 75: 180e187. https://doi.org/10.1016/j.ejsobi.2016.06.001
Ribera, C., Mas, E.D. 2015. Description of three new troglobiontic species of Cybaeodes
(Araneae, Liocranidae) endemic to the Iberian Peninsula. Zootaxa
3957(3): 313-323. https://doi.org/10.11646/zootaxa.3957.3.4
Ribera, I., Cieslak, A., Faille, A., Fresneda, J. 2018. Historical and Ecological Factors Determining Cave Diversity. En:
Moldovan, O., Kováč, Ľ., Halse, S. (eds.). Cave Ecology. Ecological Studies, pp. 229–252. Springer, Cham, Switzerland. https://doi.org/10.1007/978-3-319-98852-8_10
Růžička, V. 1989. Spider (Araneae) communities of rock debris on a typical
hillside in the in the Ceske Stredohori Mountains (North Bohemia). Acta
Entomologica Bohemoslovaca 86(6): 419-431.
Schiner, J.R. 1854. Fauna der Adelsberger-Luegr. Und Magdalenen Grotte. En: A. Schmidt (Ed.), Die Grotten und Hohlen von Adelsberg, Lueg,
Planina und Loos, pp. 231-272. Braumuller, Wien, Cit. apud.
Racovitza (1907), 435 p.
Sendra, A., Jiménez-Valverde, A., Gilgado, J.D., Ledesma, E.,
Baquero, E., Pérez-Suárez, G., Cuesta, E., et al. 2017. Diplurans of subsurface terrestrial habitats in
the Iberian Peninsula, with a new species description (Diplura: Campodeidae). Zootaxa
4291(1): 61-80. https://doi.org/10.11646/zootaxa.4291.1.4
Sket, B. 2008. Can we agree on an ecological classification of
subterranean animals? Journal of Natural History 42(21-22): 1549-1563. https://doi.org/10.1080/00222930801995762
Turner, D.D. 2019. In defense of living fossils. Biology &
Philosophy 34(2): 23. https://doi.org/10.1007/s10539-019-9678-y
Uéno, S.-I. 1980. The anophthalmic trechine beetles of the group of Trechiama
ohshimai. Bulletin of the National Science Museum, Tokyo, Serie A,
6(4): 195-274.
Uéno, S.-I. 1981. New anophthalmic Trechiama (Coleoptera,
Trechinae) from northern Shikoku, Japan. Journal of the speleological
Society of Japan 6: 11-18.
Vandel, A. 1965. Biospéologie: La Biologie des Animaux cavernicoles. Ed.
Gauthier-Villars, Paris, France.
Viré, A. 1900. La
faune souterraine de France. Paris, France.