Introducción
Los ecosistemas están sujetos a
perturbaciones naturales y antrópicas que alteran la estructura y función de
sus comunidades biológicas y los servicios ecosistémicos que nos ofrecen (IPBES 2019). Las sequías, las inundaciones, los incendios
o los tornados son ejemplos de perturbaciones naturales, mientras que los
cambios en el uso del suelo, el cambio climático o las invasiones biológicas
son algunas de las perturbaciones antrópicas más comunes (IPCC
2022). Ambos tipos de perturbaciones han incrementado en frecuencia e
intensidad durante las últimas décadas por el impacto creciente del cambio
global y a menudo se superponen sinérgicamente, desafiando las acciones de
conservación y gestión. Un ejemplo de estos ecosistemas son los ríos temporales
(RTs), definidos como ecosistemas fluviales que dejan de fluir en algún momento
del año o que se secan por completo. En estos ríos, la temporalidad del flujo o
del agua puede considerarse una perturbación natural a la que se le añaden las
perturbaciones antrópicas. Los RTs representan aproximadamente el 60% de la red
fluvial mundial y son los ecosistemas fluviales dominantes en las regiones
áridas y semiáridas, como el Mediterráneo (Messager
et al. 2021). Además, las proyecciones futuras sugieren que su extensión,
en el espacio y en el tiempo, aumentará significativamente en algunas regiones
como resultado de los cambios en el clima, el uso del suelo, o su interacción (Döll y Schmied 2012). A pesar de su relevancia
mundial y de los numerosos servicios ecosistémicos que proporcionan (Datry et al. 2018; Vidal-Abarca
et al. 2023), los RTs han sido históricamente ignorados por investigadores,
gestores y la sociedad en general (Acuña et al. 2014).
En las últimas décadas se ha avanzado de manera significativa en su
conocimiento, pero todavía persisten muchas brechas científicas y de gestión (Datry et al. 2021).
Los cambios hidrológicos de los RTs permiten la presencia de
diferentes estados acuáticos que difieren en la proporción de los diferentes
hábitats disponibles. Gallart y colaboradores (2012) proponen categorizar la dinámica temporal de los RTs en seis estados acuáticos
útiles para la gestión de estos ríos (Fig. 1). La
duración de cada uno de ellos depende del régimen de precipitación anual y de
las condiciones geomorfológicas locales, tales como el tipo y permeabilidad del
sustrato (Bonada et al. 2007). A menudo, estos
estados acuáticos se agrupan en tres fases: fluyente, de pozas y seca. La fase
fluyente está caracterizada por la presencia de caudal circulante e incluye
rápidos (hábitats lóticos), más o menos predominantes, conectados a pozas
(hábitats lénticos). La fase de pozas incluye, exclusivamente, pozas
desconectadas (hábitats lénticos), mientras que la fase seca incluye cauces
totalmente secos (hábitats terrestres) (Gallart et al.
2012) (Fig. 1).

Figura 1. Estados acuáticos según Gallart et al. (2012) agrupados según si corresponden
a la fase fluyente, de pozas desconectadas o a la fase de cauces secos. Créditos: MIRAGE and TRivers projects.
Figure
1. Aquatic states according to Gallart et al. (2012) grouped based on whether they correspond to the flowing phase,
disconnected pools phase, or dry riverbed phase. Photo credits: MIRAGE and
TRivers projects.
Las especies, tanto acuáticas como terrestres, se alternan
entre los tres tipos de fases, lo que resulta en una elevada diversidad beta
temporal (Tonkin et al. 2017; Sánchez-Montoya et al. 2020a, 2022). Sin embargo, desde una perspectiva
acuática, los RTs tienen menor diversidad local (diversidad alfa) que los ríos
permanentes (Soria et al. 2017), y ésta disminuye
significativamente al aumentar la duración de la fase seca (Datry et al. 2014; Sánchez-Montoya
et al. 2018). La pérdida de caudal implica la desaparición de especies
acuáticas adaptadas a los hábitats lóticos, pero también la aparición de
especies que se encuentran exclusivamente en las pozas desconectadas. Con el
tiempo, las condiciones abióticas de las pozas cambian y, con ellas, la
composición de especies de sus comunidades acuáticas, dependiendo de cuándo se
hayan desconectado las pozas y de las características locales (Bonada et al. 2020). Estas pozas pueden albergar
muchas especies y contribuir significativamente a la biodiversidad de algunos
RTs, o pueden tener una diversidad muy pobre debido a la adversidad de las
condiciones abióticas (p.e. concentración baja de oxígeno disuelto en el agua; Bonada et al. 2020). Eventualmente, la pérdida de agua
superficial desencadena respuestas variadas. Algunos organismos acuáticos
simplemente mueren; otros emergen y completan su ciclo en el ecosistema
terrestre; otros migran a otras pozas o al hiporreos; otros pueden mantenerse
activos mientras haya humedad (p.e. los microorganismos; Arce
et al. 2019; Vidal-Abarca et al. 2020); y
otros activan formas de resistencia que permanecen latentes en el cauce seco
hasta la reanudación del caudal después de la lluvia (Bogan
et al. 2017). Esta reanudación del caudal es otro momento clave para las
comunidades acuáticas. Los organismos recolonizan los hábitats que ahora fluyen
y las comunidades se reorganizan siguiendo procesos de sucesión secundaria (Bonada et al. 2020) (Fig. 2).
Desde una perspectiva terrestre, a medida que el flujo de agua superficial comienza a desaparecer, los organismos terrestres colonizan los cauces secos desde los ecosistemas adyacentes (riberas y
laderas: Steward et al. 2022; Sánchez-Montoya et al. 2016) (Fig. 2). Estas comunidades terrestres son clave desde una
perspectiva taxonómica, funcional y
ecológica (Sánchez-Montoya et al. 2020a, 2020b), pero han sido mucho menos estudiadas que las
comunidades acuáticas. Los estudios existentes en Europa (Francia: Corti y Datry 2016;
España: Sánchez-Montoya et al. 2016, 2020b; Vidal-Abarca et al.
2023) revelan que los cauces secos soportan una
elevada diversidad de invertebrados y plantas terrestres, que podría ser
altamente dependiente de la humedad del sedimento y cómo ésta se preserve a lo
largo de la fase seca (Sánchez-Montoya
et al. 2016).
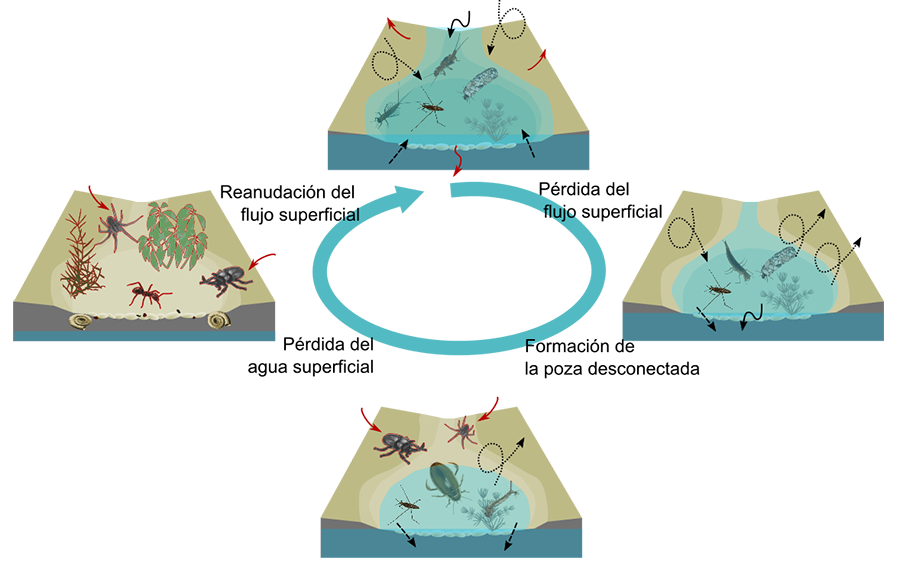
Figura 2. Cambios en
la composición de especies entre comunidades acuáticas y terrestres a lo largo
del ciclo hidrológico de los ríos temporales. Las flechas negras continuas
indican organismos que llegan o salen a través de la vía fluvial, las flechas
con puntos indican organismos que llegan o salen por vía aérea, mientras que
las flechas con líneas discontinuas indican organismos que recolonizan a partir
de formas de resistencia y que estaban en el hiporreos. Los hábitats terrestres
que aparecen con el secado están colonizados (flechas rojas) por diferentes
especies de plantas e invertebrados terrestres (símbolos con borde rojo).
Figure 2. Changes in species composition between aquatic and terrestrial
communities throughout the hydrological cycle of temporary rivers. Solid black
arrows indicate organisms that arrive or depart through the waterway, dotted
arrows indicate organisms that arrive or depart through the air, while dashed
arrows indicate organisms that recolonize from resistant forms remaining in the
hyporheic zone. Terrestrial habitats that appear during drying are colonized
(red arrows) by different species of terrestrial plants and invertebrates
(symbols with red stroke).
Las
comunidades de invertebrados terrestres están constituidas por hormigas,
colémbolos y ácaros, como los más numerosos en los ámbitos mediterráneos, junto
a arañas y escarabajos, importantes por su contribución a la diversidad
faunística, frecuencia y biomasa. También destacan crustáceos isópodos y
diversos insectos (como grillos y tijeretas, algunos propios de estos
ambientes), sin olvidar gasterópodos (caracoles y babosas) y otros
invertebrados no artrópodos (lombrices de tierra u oligoquetos). En formas más
edáficas de los cauces secos aparecen otros grupos complementarios en la
comunidad, tales como los miriápodos (quilópodos o ciempiés de varios órdenes y
diplópodos o milpiés), o grupos menos conocidos de insectos como los
psocópteros, embiópteros, o insectos sin alas como zigentomos y
microcorifios (pececillos plateados y pececillos
saltarines, respectivamente) (Sánchez-Montoya
et al. 2016, 2020b; Steward et
al. 2017, 2022). En cuanto a las comunidades
vegetales terrestres, las especies anuales reemplazan gradualmente a las
acuáticas durante la fase seca, y los cauces secos
que exhiben condiciones secas prolongadas pueden colonizarse completamente por
plantas terrestres perennes (Vidal-Abarca
et al. 2020; Martínez-Yoshino et al. 2021). Estas plantas influyen en su entorno fluvial, actuando
como ingenieros del ecosistema al facilitar la acumulación de sedimentos con
nutrientes y material orgánico que estarán disponibles para los organismos río
abajo, con la reanudación del caudal (Steward
et al. 2012).
La Directiva Marco del Agua (DMA; Comisión Europea 2000) representó un cambio de
paradigma y una revolución en la gestión de los ecosistemas fluviales (Moss 2008). Sin embargo, a pesar de estos avances, la DMA
no considera explícitamente los RTs. Sí se incluyeron, en parte, en el proceso
de intercalibración entre los estados miembros mediterráneos con el fin de
garantizar la comparabilidad entre los métodos y las clases de calidad
desarrollados por cada país (Comisión Europea
2011), pero en la actualidad, cada estado miembro utiliza sus propios
métodos para evaluar el estado ecológico de los RTs (Feio
et al. 2014; Reyjol et al. 2014), o simplemente
no son considerados como masa de agua (p.e. en Francia: D5.2 en www.dryver.eu/results/reports-and-documents). En España, los RTs se incorporaron de
manera preliminar en la normativa legal relativa a la evaluación del estado de
las masas de agua desarrollada para la implementación de la DMA mediante la
Instrucción de Planificación Hidrológica (IPH) (Orden ARM/2656/2008, de 10 de
septiembre (MARM 2008)), y posteriormente mediante el Real
Decreto 817/2015 de evaluación del estado de las aguas superficiales (MAGRAMA 2015). En la IPH se mencionan específicamente
a los RTs para el establecimiento de los caudales ambientales y para la
evaluación hidrológica, donde los ríos se clasifican en 4 tipologías:
permanentes, temporales o estacionales, intermitentes o fuertemente estacionales,
y efímeros. Estas tipologías se definieron en base al número de días con
condiciones de flujo al año, pero esta clasificación no se trasladó a los tipos
de ríos para la evaluación ecológica. Posteriormente, el Real Decreto define 37
tipos de ríos, con las correspondientes condiciones de referencia y límites de
cambio de clases de calidad. Tres de estos tipos se refieren a los RTs de las
Islas Baleares, demarcación hidrográfica en la que todos los ríos son
temporales, que fueron incorporados en el Plan Hidrológico de las Islas
Baleares del año 2009 y desarrollados en el del año 2011 (García y Rodríguez-Lozano 2020). Ninguno
de los otros tipos para el resto de las demarcaciones hidrográficas se refiere
a RTs, aunque la mayoría de los tipos definidos contienen una elevada
proporción de RTs. Más recientemente, y en base a los estudios previos de Gallart et al. (2012), en las cuencas catalanas (NE
España) se han incorporado en el Plan de Cuenca tipologías adicionales de RTs y
un protocolo para su clasificación y evaluación de su estado ecológico (Munné et al. 2021).
Según
la DMA, el estado ecológico de los ríos es una medida de calidad definida a
partir de elementos biológicos, hidrogeomorfológicos y fisicoquímicos (Comisión Europea
2000). La evaluación de la
calidad biológica considera varios elementos biológicos (diatomeas, macrófitos,
macroinvertebrados y peces) y se determina en relación con un tipo de río
establecido y su condición de referencia (Bailey et al. 2004). Esta calidad biológica se complementa con
elementos de calidad hidrogeomorfológica (p.e., caudal, calidad de la
vegetación de ribera, o aspectos sobre la morfología fluvial; Florsheim et al.
2008; Ollero et al. 2021), la fisicoquímica general (p.e., oxígeno y
nutrientes), y la concentración de contaminantes específicos (incluyendo las
sustancias preferentes, es decir, aquellas que presentan un riesgo
significativo para las aguas superficiales debido a su especial toxicidad,
persistencia y bioacumulación) (p. e., Tueros et al. 2009). Sin embargo, las metodologías actuales para
evaluar el estado ecológico fueron diseñadas y desarrolladas para ríos
permanentes y resultan en una inexacta evaluación del estado ecológico de los
RTs (Tabla 1). Esto se debe a que no permiten distinguir entre
los efectos de las perturbaciones naturales (temporalidad del agua) de las
antropogénicas (Soria
et al. 2020), muy probablemente
porque las mismas características de resiliencia y resistencia que permiten a
las especies hacer frente a la temporalidad, les permiten hacer frente a las perturbaciones
antrópicas (Vinebrooke et al. 2004).
Por ejemplo, los índices de macroinvertebrados estandarizados aplicados a RTs
de referencia no consiguen llegar a los niveles de calidad que les corresponde,
incluso considerando niveles de calidad adaptados para estos ríos (Munné et al. 2021) (Tabla 1). Para macrófitos, en España, sólo el 52% de los tramos de referencia
que son RTs (n=25) alcanzan el “Muy buen estado”, mientras que para los
permanentes (n=202) este porcentaje sería del 84% (Mellado-Díaz, com. pers.).
Tabla 1. Índices
biológicos más utilizados en España para la evaluación del estado ecológico de
las masas de agua según la Directiva Marco del Agua (DMA) y funcionamiento en
ríos temporales (RTs).
Table
1. Most commonly used biological indicators in
Spain for assessing the ecological status of water bodies according to the
Water Framework Directive (WFD) and their performance in temporary rivers
(RTs).
Debido al limitado funcionamiento de los índices biológicos
en los RTs, en los últimos años en España se han empezado a diseñar
metodologías específicas para estos ríos. Por ejemplo, en el caso de la
Demarcación Hidrográfica Islas Baleares, en la que todos los ríos son RTs, se
han desarrollado índices biológicos para diatomeas y macroinvertebrados (Delgado et al. 2012; García
et al. 2014). Por su parte, en el marco del proyecto LIFE+ TRivers se desarrolló el TREHS (Estado Ecológico e Hidrológico de los Ríos Temporales)
(Gallart et al. 2017), una herramienta que permite
a los gestores (1) conocer el régimen hidrológico real de los RTs incluso sin
datos de estaciones de aforo, (2) evaluar el estado hidrológico de un río
comparando el régimen hidrológico real con el natural, (3) proporcionar un
calendario sobre el estado acuático más probable en cada momento para guiar el
muestreo, y (4) clasificar y representar en un triángulo denominado FPD (Flow,
Pools and Dry plot, por sus siglas en inglés) los RTs en 8 hidrotipos, en
función de la frecuencia relativa de flujo (Mf), pozas desconectadas (Mp) y
cauces secos (Md) a lo largo de un año (Fig. 3a).
Más recientemente, la “Guía para la evaluación de las aguas superficiales y
subterráneas”, publicada en
octubre de 2020 (MITECO 2020 —ver Anexo 1 de la guía—),
adopta la metodología TREHS y simplifica los 8 hidrotipos en los 4 para
caudales ambientales definidos en la IPH. Además, adapta los valores de Mf, Mp
y Md para estos 4 hidrotipos y sugiere estrategias de muestreo específicas para
cada uno (Fig. 3b, c). Así, según esta guía, el
primer hidrotipo considera las masas de agua permanentes
y cuasipermanentes (H1: H1-1 y H1-2, respectivamente), que deben evaluarse sin
modificación de los protocolos. El segundo hidrotipo (H2) considera los RTs que
fluyen durante una gran parte del año, y que deben ser evaluados adaptando el
calendario de muestreo. El tercer hidrotipo (H3) incluye las masas de agua con
pozas desconectadas durante una gran parte del año, y para los que todavía se
necesitan desarrollar nuevos índices que permitan una evaluación correcta de la
calidad biológica. Finalmente, el cuarto hidrotipo (H4) considera conjuntamente
los RTs ocasionales y episódicos, que están secos durante una gran parte del
año. Para este último es posible que las comunidades biológicas (acuáticas) no
tengan tiempo suficiente para establecerse por completo, y se sugiere que se
recurra a índices exclusivamente hidrogeomorfológicos, a pesar de que las
comunidades terrestres podrían ser una buena alternativa si se desarrollan indicadores
específicos (Stubbington et al. 2019).
Las características de los RTs y las evidencias existentes
hasta la fecha ponen de manifiesto numerosas lagunas de conocimiento que
limitan el desarrollo de metodologías adecuadas para la evaluación de su estado
ecológico. Así pues, el objetivo de esta revisión es identificar los retos
actuales para la correcta evaluación del estado ecológico de los RTs y analizar
las oportunidades existentes para hacer frente a estos retos. La revisión se
centra mayoritariamente en el conocimiento científico y de gestión acumulado
desde la implementación de la DMA en España, pero recoge experiencias en otros
RTs del mundo para guiar acciones de conservación y gestión en estos
ecosistemas únicos y altamente amenazados por el cambio global.
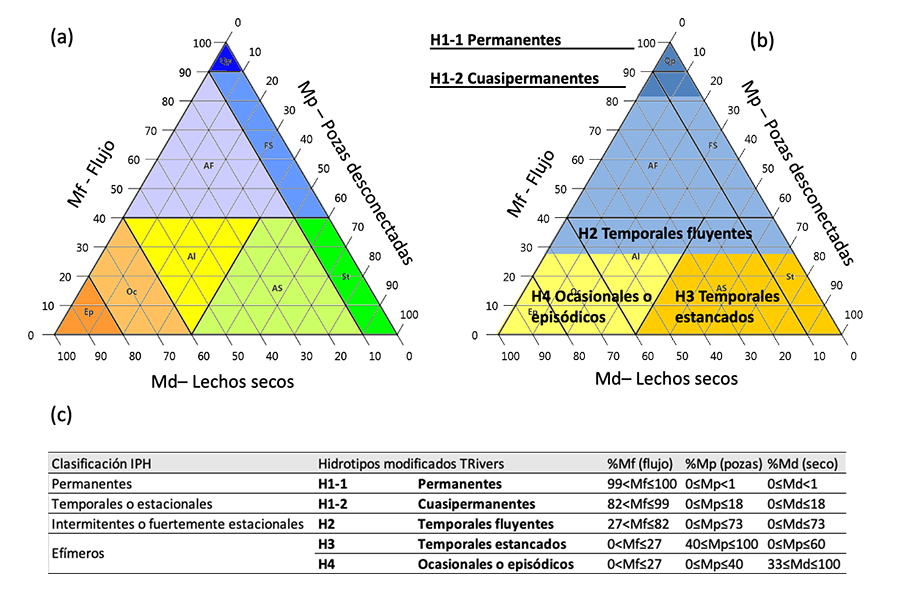
Figura 3. (a) Triángulo FPD del software TREHS
desarrollado en el proyecto LIFE+ TRivers que
identifica 8 hidrotipos (incluidos los permanentes) para ríos temporales (Gallart et al. 2017) en función de las frecuencias de
las tres fases (%): Qp para cuasipermanentes, AF para fluyentes alternos, FS
para fluyentes estancados, St para estancados, AS para alternos estancados, Al
para alternos, Oc para ocasionales y Ep para episódicos; (b) adaptación
del triángulo FPD a los 4 hidrotipos para caudales ambientales definidos en la
IPH (Instrucción de Planificación Hidrológica), y (c) frecuencia de
caudal, pozas desconectadas y cauces secos para los 4 hidrotipos de la IPH y su
equivalencia en el proyecto TRivers (b). Adaptado del Anexo 1 de la
“Guía para la evaluación de las aguas superficiales y subterráneas” (MITECO 2020).
Figure 3. (a) FPD triangle from the TREHS software developed in the LIFE+
TRivers, identifying 8 hydrotypes (including
permanent ones) for temporary rivers (Gallart et al. 2017) using the
frequencies of the three phases (%): Qp for quasi-permanent, AF for alternating
flow, FS for stagnant flow, St for stagnant, AS for alternating stagnant, Al
for alternating, Oc for occasional, and Ep for episodic; (b) adaptation of the FPD triangle to
the 4 environmental flow hydrotypes defined in the IPH (Hydrological Planning
Instruction), and (c)
flow frequency, disconnected pools, and dry channels for the 4 IPH hydrotypes
and their equivalence in the TRivers project (b). Adapted from Annex 1 of the "Guía para la evaluación
de las aguas superficiales y subterráneas” (MITECO 2020).
Retos para la evaluación del estado ecológico en ríos
temporales
RETO 1:
Diferenciación entre ríos temporales naturales o hidrológicamente impactados
A pesar de que la temporalidad es un fenómeno natural en los
ríos de clima Mediterráneo, algunos RTs son ríos permanentes hidrológicamente
impactados, cuyo carácter temporal es consecuencia de la actividad humana (p.e.
extracciones de agua; Chiu et al. 2017). Asimismo,
aunque menos común, algunos RTs naturales se han transformado en permanentes
tras recibir efluentes de depuradora o excedentes de canales de riego (Martín-Vide 1999). Diferenciar entre RTs naturales
o hidrológicamente impactados es complejo, y disponer de información sobre el
régimen natural del río, así como de sus impactos hidrológicos potenciales es
clave.
La información sobre el régimen natural del río se puede
obtener a partir de modelos de precipitación-escorrentía, de registros
obtenidos antes de su alteración, o de registros de ríos en condiciones
climáticas y fisiográficas similares. Cuando se dispone de la información
adecuada del régimen natural y del real el grado de alteración hidrológica se
suele estimar comparando diversas métricas, cuya influencia sobre la vida
acuática se conoce o supone (Poff et al. 2010).
Normalmente las métricas utilizadas para estimar la alteración hidrológica de
los ríos se basan en variables que consideran el régimen de caudales, lo cual
es muy complejo de obtener para RTs. Por lo tanto, es necesario el desarrollo
de métricas alternativas que consideren datos complementarios (Gallart et al. 2017). Dentro de las herramientas
disponibles en la actualidad, la herramienta TREHS permite estimar 6 métricas
del régimen temporal de los ríos temporales utilizando diversos tipos de
información, y estimar el grado de alteración hidrológica en base a unas tablas
de ‘criterio de experto’ que pueden ser modificadas por el usuario si éste
dispone de la información adecuada para valorar la relevancia de cada métrica
sobre la vida acuática. Por otra parte, la herramienta BIOAS-Tool (Cid et al. 2016) permite deducir el estado hidrológico a
partir de las comunidades biológicas, y éste se podría contrastar con la
alteración hidrológica de la cuenca (es decir, impactos que puedan
potencialmente modificar el régimen hidrológicos como extracciones, presas o
azudes). Algo similar fue propuesto por Fritz y colaboradores en EEUU para
inferir la permanencia del flujo utilizando musgos acuáticos y ribereños (Fritz et al. 2009), o más recientemente,
macroinvertebrados acuáticos (Fritz et al. 2023).
RETO 2:
Diferenciación entre perturbación natural y antrópica
Existen especies con adaptaciones específicas a los RTs (Bogan et al. 2017) pero, en general, las comunidades
acuáticas de estos ríos están formadas por pocas especies de carácter
generalista, lo que les permite hacer frente a las condiciones cambiantes. Ello
conlleva que la respuesta a las perturbaciones antrópicas no sea clara y pueda
confundirse con los efectos de la propia perturbación natural que supone la
temporalidad, más aún cuando los dos tipos de perturbaciones actúan
simultáneamente (Bonada y Resh 2013; Belmar et al. 2019; Soria et
al. 2020). Por ejemplo, en las pozas desconectadas de los RTs pueden
acumularse grandes cantidades de detritus que pueden eutrofizar el agua, efecto
similar al que podría generarse a causa de la contaminación por fertilizantes o
aguas residuales (Bonada et al. 2020). Además, y
teniendo en cuenta la elevada variabilidad espacial y temporal de los RTs (p.e.
las comunidades cambian entre estados acuáticos, en función de la duración del
periodo de sequía), las comunidades acuáticas son, de manera natural, muy
variables (Bonada et al. 2020). Estas
circunstancias hacen que definir una comunidad de referencia para cada
hidrotipo sea complejo.
Estudios previos han mostrado que existen taxones (p.e.
Hydrophilidae) cuya presencia durante la fase de flujo ya indicaría que se
trata de un RT de referencia sin tener que disponer de datos hidrológicos (Cid et al. 2016). Ello supone un primer paso para la
determinación de las condiciones de referencia en RTs, pero la gran
variabilidad de las comunidades de estos ríos limita su generalización, con lo
que se generan dudas de si realmente es posible definir condiciones de
referencia fijas para RTs o si éstas deberían ser cambiantes en función del
estado acuático en el que se encuentran, además de incorporar información a
largo plazo sobre su variabilidad interanual (Soria et
al. 2020). De hecho, hay estudios que muestran cómo los índices de calidad
biológica actuales sólo funcionan en los RTs si se trata de un año húmedo (Munné y Prat 2011; Mazor et
al. 2014) y si la permanencia del flujo es lo suficientemente larga y
predecible (p.e. hidrotipos cuasipermanentes o temporales fluyentes; Gallart et al. 2012, 2017).
Otra opción sería utilizar nuevos índices de calidad biológica a nivel de
género o especie, o índices funcionales basados en rasgos biológicos
específicos que puedan responder a las perturbaciones antrópicas
independientemente de la temporalidad (Soria et al. 2020).
RETO 3:
Desarrollo de índices biológicos para pozas desconectadas
La importancia de considerar las
pozas desconectadas como parte de la evaluación del estado ecológico de una
masa de agua se fundamenta en que: 1) todas las masas de agua con algún grado
de temporalidad pueden presentar pozas entre el 18 y el 100 % del tiempo a
escala anual (ver Fig. 3c); 2) las administraciones competentes en materia de
gestión del Dominio Público Hidráulico actualmente no evalúan el estado
ecológico de las pozas desconectadas por falta de metodologías apropiadas; 3)
las pozas son hábitats clave para la conservación de numerosas especies, ya que
actúan como refugios de biodiversidad acuática a nivel local y regional; 4) las
pozas son hábitats transitorios para especies lóticas (durante las etapas
iniciales) y lénticas; 5) las pozas pueden tener intercambios limitados o nulos
con aguas subterráneas cuya calidad fisicoquímica se altera de manera natural;
y, finalmente, 6) se espera que la presencia de pozas en el cauce de los ríos
aumente de la misma manera que aumentarán los RTs por el cambio global (Bonada et al. 2020) (Fig. 4).
En la actualidad, no existen
índices diseñados específicamente para evaluar la calidad biológica de las
pozas desconectadas, y los que se usan en ríos permanentes o que pueden
funcionar en RTs, no se aplican porque algunas de estas pozas pueden tener
pocas especies de manera natural, y la mayoría son tolerantes a la
contaminación (Bonada et al. 2020). Los índices para estas pozas podrían considerar
diatomeas, macroinvertebrados u otros elementos de calidad utilizados en
ecosistemas lénticos (p.e. anfibios o
comunidades planctónicas; Van den Broeck
et al. 2015). Los macrófitos también pueden ser
comunes en pozas desconectadas pero su presencia es irregular y, por lo tanto,
hay menos oportunidades de utilizarlos como elemento biológico. No obstante,
los macrófitos han sido propuestos como bioindicadores en pequeños ecosistemas
lénticos (p.ej. Labat et al. 2022), por lo que sería posible la adaptación de alguno de
estos índices o metodologías a las particularidades de las pozas desconectadas.
Los peces son elementos clave desde el punto de vista de la conservación en
pozas desconectadas, pero a menudo no están presentes o, si lo están, las
comunidades están formadas por una o pocas especies (Bonada et al. 2020).
Asimismo, los índices funcionales basados en rasgos biológicos (p.e. redundancia funcional de la comunidad de
macroinvertebrados) también podrían ser una buena opción para pozas
desconectadas (Soria et al. 2020), aunque éstos métodos están todavía en fases muy
preliminares, y son necesarias más investigaciones para elaborar índices
alternativos y estandarizar sus puntuaciones para poder hacer comparaciones
entre las cuencas hidrográficas mediterráneas (p.e. Magalhaes et al. 2008 o
Hermoso et al. 2010).
El desarrollo de índices biológicos específicos
para pozas desconectadas requeriría considerar
tanto los factores locales como regionales que determinan
las comunidades biológicas. Los factores locales incluirían el régimen hidrológico
(p.e. pozas
confinadas por roca madre frente a pozas con interacciones de agua subterránea;
Bonada et al. 2020),
la fisicoquímica del agua, el tamaño de la poza, la heterogeneidad del
sustrato, el tiempo transcurrido desde la desconexión, la duración de la poza y las interacciones bióticas que puedan establecerse. Los
factores regionales incluirían la distancia a
otros hábitats de agua dulce que actúan como fuente de organismos, incluidas
otras pozas desconectadas o masas de agua permanentes de la misma cuenca
o de cuencas cercanas (Cid et al. 2020).

Figura 4. Pozas desconectadas y cauces secos de
ríos temporales de la península ibérica. (a) Río Despeñaperros (cuenca del
Guadalquivir); (b) Río Algars (cuenca del Ebro); (c) Riera de Rellinars (cuenca
del Llobregat); (d) Río Daró (cuenca del Daró); (e) Barranc de la Fou (cuenca
del Júcar); (f) Rambla del Carcelín (cuenca del Segura); (g) Arroyo Arjona y
Migo López (cuenca del Guadalquivir); (h) Riera de Gotarra (cuenca del Ter);
(i) Rambla de Espinardo (cuenca del Segura); (j) Barranc de Fou (cuenca del Sènia).
Figure
4. Disconnected pools and dry riverbeds of
temporary rivers in the Iberian Peninsula. (a) Despeñaperros River
(Guadalquivir basin); (b) Algars River (Ebro basin); (c) Riera de Rellinars
(Llobregat basin); (d) Daró River (Daró basin); (e) Barranc de la Fou (Júcar
basin); (f) Rambla del Carcelín (Segura basin); (g) Arroyo Arjona and Migo López
(Guadalquivir basin); (h) Riera de Gotarra (Ter basin); (i) Rambla de Espinardo
(Segura basin); (j) Barranc de Fou (Sènia basin).
RETO 4: Desarrollo de
índices biológicos para cauces secos
La fase de flujo es corta en ríos ocasionales y episódicos
(por debajo del 27% del año, Fig. 3c y
4), resultando en comunidades acuáticas con
pocas especies, la mayoría generalistas, ya que el secado comienza antes de que
se desarrollen comunidades complejas y diversas. Por lo tanto, no se prevé que
el desarrollo de herramientas basadas en elementos acuáticos sea útil, y existe
una necesidad urgente de identificar nuevos indicadores que, por ejemplo,
consideren las comunidades biológicas terrestres de cauces secos (Steward et al. 2018; Stubbington
et al. 2019).
Los invertebrados terrestres empiezan a ser reconocidos como
buenos bioindicadores en ecosistemas terrestres adyacentes a ríos como son las
riberas (Hodkinson y Jackson 2005), pero, a
diferencia de los macroinvertebrados acuáticos, casi no existe conocimiento
científico sobre su tolerancia ambiental a diferentes impactos (con la
excepción de Steward et al. 2018), por lo que a la
fecha no se han considerado en la evaluación rutinaria del estado ecológico (Stubbington et al. 2019). El escaso
reconocimiento de los indicadores de invertebrados terrestres está relacionado
con la inercia histórica en la investigación compartimentada (acuática vs.
terrestre) y la taxonomía relativamente compleja de estos organismos. Steward et al. (2018) encontraron que los
invertebrados terrestres responden a la perturbación física del sustrato por
parte del ganado y los mamíferos salvajes en los ríos australianos secos y, por
lo tanto, pueden ser indicadores efectivos de la calidad de la fase seca. Por
lo tanto, los invertebrados terrestres cumplen potencialmente con la mayoría de
los criterios de bioindicadores adecuados ya que (1) son abundantes y comunes
en cauces de ríos secos en todo el mundo, (2) son fáciles de muestrear y
consistentes con equipos de muestreo económicos, (3) son sensibles a las
perturbaciones antrópicas en los hábitats terrestres (bosques, zonas ribereñas
y matorrales) (Gerlach et al. 2013), y (4) su
naturaleza sedentaria básica, en especial de aquellos que habitan en el suelo,
facilita el análisis espacial de los efectos de la contaminación.
Las plantas terrestres se han utilizado en gran medida como
indicadores de la salud de los ríos, pero solo considerando los macrófitos como
elemento de la calidad biológica o las plantas de ribera como elemento de la
calidad hidrogeomorfológica (p.e. QBR; Munné et al. 2003).
Sin embargo, la estrecha interacción entre las plantas y su entorno las
convierte en indicadores potencialmente útiles de las diferentes condiciones
del hábitat, incluidas las resultantes de la intermitencia del flujo (Westwood et al. 2021). Por otro lado, a pesar de su
dependencia del agua, las macroalgas y los briófitos también prosperan en
sistemas temporales, donde han desarrollado formas de vida y estrategias para
hacer frente a la estacionalidad del flujo y del agua (Vieira
et al. 2018). Recientemente, se ha demostrado que las plantas que colonizan
los cauces secos de los ríos responden a impactos humanos no hidrológicos, como
la composición de los sedimentos, el sombreado y los impactos geomorfológicos (Stubbington et al. 2019), lo que indica que
también tienen un alto potencial como bioindicadores. Sin embargo, es necesario
llenar muchos vacíos en la investigación antes de que las plantas se utilicen
de forma rutinaria en la evaluación del estado ecológico (Westwood et al. 2021).
RETO 5: Evaluación de la calidad mediante el uso de
indicadores físico-químicos
Conocer qué métricas
fisicoquímicas habría que utilizar en la evaluación del estado ecológico de los
RTs también representa un reto, ya que muchas de ellas se ven alteradas de
manera natural, incluidas las que se utilizan en ecosistemas lénticos (Gómez et al. 2017). Por ejemplo, la disponibilidad del fósforo en
el sedimento podría ser una buena medida de la eutrofización en RTs, como lo es
en humedales (Mesnage
et al. 2002), pero los nutrientes
pueden ser significativamente altos de forma natural en las pozas
desconectadas, debido a la evaporación del agua o, en condiciones de anoxia,
por mineralización de la materia orgánica y/o ausencia de recirculación (Fellman et al. 2011). De manera similar, la concentración de oxígeno
disuelto puede experimentar una alta variabilidad diaria e incluso agotarse
hasta la anoxia, lo que puede movilizar nutrientes de los sedimentos y
disminuir la calidad química de la poza (Gómez et al. 2017). La evaporación de las pozas desconectadas
también controla la salinidad, especialmente cuando las pozas están conectadas
a aguas subterráneas (Gómez et al. 2017). Por lo
tanto, al evaluar la calidad fisicoquímica general de las pozas desconectadas,
se deberá tener en cuenta que las alteraciones de estos indicadores
fisicoquímicos no necesariamente reflejan la presencia de impacto humano, sino
también las características naturales de estos hábitats.
Además de los elementos biológicos, la fisicoquímica de los
sedimentos secos también podría contribuir a la evaluación del estado ecológico
de los ríos ocasionales y episódicos. Los cauces de ríos secos son áreas de
almacenamiento de nutrientes y material orgánico, de procedencia tanto local
como de toda la cuenca (Steward et al. 2012). De
hecho, algunos parámetros fisicoquímicos (p.e. conductividad, contenido de
nutrientes, pH y contenido de carbono) en los sedimentos del cauce seco de los
ríos son potencialmente buenos indicadores de la calidad fisicoquímica durante
la fase seca (Sánchez-Montoya y Arce, com. pers.). Por ejemplo, el contenido en
nitrato de los sedimentos secos de un tramo parece ser sensible a la intensidad
de agricultura de regadío presente en la cuenca (Arce et
al. 2023). Además, la cantidad y forma de los nutrientes en los sedimentos
del cauce del río también representan una carga interna potencial que puede
liberarse parcialmente durante la reanudación del flujo y condicionar la
fisico-química en la fase fluyente (Arce et al. 2014).
Un caso claro de impacto donde los elementos físico-químicos
podrían actuar como indicadores puede verse en aquellos RTs que actúan como
receptores directos de efluentes urbanos o industriales, impactando
adicionalmente el sistema con el desarrollo puntual de cierto caudal (Skoulikidis 2008; Mandaric
et al. 2019). En estos casos, el análisis de parámetros químicos
específicos de la actividad, como por ejemplo contaminantes orgánicos,
sustancias farmacéuticas o metales pesados, debe ser considerado incluso en los
sedimentos a pesar del cesado del efluente. Sin embargo, la definición de
estándares de calidad, así como la aplicabilidad práctica de estos parámetros
en la evaluación rutinaria del estado ecológico, debe investigarse más a fondo.
RETO 6:
Adaptación de los índices hidrogeomorfológicos
Los RTs, y especialmente en aquellos con pozas o cauces
secos, son el sustrato físico sobre el que se desarrollan distintos y complejos
procesos hidrogeoquímicos y biológicos (Arce et al. 2019).
La dinámica geomorfológica de un río y el funcionamiento de los procesos
físicos naturales generan espontáneamente la creación y el mantenimiento de
hábitats y aseguran la integridad del ecosistema (p.e. Florsheim et al. 2008). Su deterioro supone la
alteración de muchos de estos procesos y, en consecuencia, contribuyen a
modificar la calidad ecológica de los RTs. Actualmente, la calidad
hidrogeomorfológica de los RTs es raramente evaluada a través de indicadores e
índices al uso, aunque existen algunos intentos propuestos por diferentes
autores: el Índice de Alteración de Ramblas (IAR: Suárez y Vidal-Abarca 2008), el Índice
Hidro-Geomorfológico adaptado a ríos efímeros (IHG: Ollero
et al. 2021), El Morphological Quality Index (MQI: Rinaldi et al. 2013), o el Geomorphic Status (GS:
Lobera et al. 2015). Estos índices tienen en cuenta
bien las alteraciones hidrológicas de los cuerpos de agua, bien las
modificaciones o impactos que sufren el cauce y el hábitat ribereño
circundante. Sin embargo, la aplicabilidad general de estos índices en los RTs
no se ha demostrado fehacientemente y, por lo tanto, todavía no se consideran
en la evaluación rutinaria del estado ecológico.
En el presente ciclo de planificación hidrológica
(2022-2027), las administraciones competentes deben aplicar el llamado
“Protocolo de caracterización hidromorfológica de masas de agua de la categoría
ríos” (Protocolo HMF), código M-R-HMF-2019 (MITECO 2019).
El Protocolo HMF intenta responder a la necesidad de incluir las condiciones
mediterráneas, y desarrolla una valoración adaptada en el caso de ríos que no
tengan capacidad de albergar fauna piscícola y vegetación de ribera,
mayoritariamente los RTs. En este sentido, resulta un avance en los métodos de
caracterización hidromorfológica oficiales, ya que se consideran
específicamente los RTs, limitación detectada en estudios comparativos previos
(Fernández et al. 2011). Sin embargo, dado el
corto período de aplicación de este Protocolo HMF y, teniendo en cuenta que
muchos RTs no están clasificados como masas de agua, no se puede evaluar aún la
conveniencia de este método y su precisión respecto a los índices anteriores.
RETO 7: Utilidad
y aplicación de la teoría de metacomunidades
Desde el desarrollo de la teoría neutral de la biodiversidad
(Hubbell 2001), el concepto de ecología de
comunidades ha cambiado significativamente nuestra visión para explicar la
composición local de especies. Las comunidades no sólo se explican por factores
ambientales deterministas locales (factores abióticos y bióticos) sino también
por factores estocásticos regionales (Rosindell
et al. 2011). Independientemente de si la dispersión es un factor
determinista o estocástico, ésta juega un papel clave para el intercambio de
individuos entre comunidades definiendo la conectividad entre ellas, hecho que
remarca su actual consideración en los estudios de ecología de comunidades (Cañedo-Argüelles et al. 2015; Sarremejane et al. 2017). Las comunidades no se
ven como entidades aisladas, sino como entidades que interactúan en el paisaje
a través de la dispersión dando lugar a metacomunidades (Leibold y Chase 2018).
La determinación del estado ecológico en ríos se ha basado,
tradicionalmente, en valoraciones locales sin considerar la perspectiva
regional, como herencia del paradigma de nicho que ha prevalecido en la
ecología durante décadas (Barton et al. 2020; Cid et al. 2020). Por ejemplo, el índice para
macroinvertebrados IBMWP se basa en la tolerancia de las familias de
macroinvertebrados a la contaminación (factor local), asumiendo que todas las
especies pueden llegar eventualmente a todos los sitios y no existen limitaciones
de dispersión: la presencia o ausencia de una familia en un sitio solo depende
de cómo sean las características locales (p. e. la físico-química) (Fig. 5). Sin embargo, el grado de aislamiento de los
puntos de muestreo o la propia capacidad de dispersión de las especies puede
impedir que éstas alcancen sus hábitats óptimos, resultando en valores de
calidad biológica más bajos de los esperados a pesar de la presencia de
hábitats favorables. Por el contrario, una alta conectividad puede favorecer el
movimiento de especies hacia hábitats subóptimos o desfavorables, aumentando
temporalmente la riqueza (Heino 2013). Ambos fenómenos
disminuyen el rendimiento de los métodos de evaluación del estado ecológico, lo
que sugiere que se deben implementar factores regionales en las metodologías de
evaluación (Fig. 5). Esto es especialmente
relevante en RTs ya que, más allá de la variación de conectividad espacial
existente en los ríos debido a la direccionalidad del flujo (tramos de aguas
abajo presentan más conectividad al recibir organismos de aguas arriba por deriva;
Brown y Swan 2010), la propia dinámica hidrológica
resulta en cambios en la conectividad a lo largo del ciclo hidrológico: desde
comunidades acuáticas conectadas durante la fase fluyente a desconectadas
durante la fase seca. Esta fuerte variabilidad espacio temporal en la
conectividad determina la diversidad y la composición de los tramos, siendo los
tramos menos conectados, espacial y temporalmente, los que presentan menor
riqueza y diferente composición (Cunillera-Montcusí
et al. 2023). Todo esto, sumado a la propia capacidad de dispersión de los
organismos, determina qué comunidades vamos a encontrar en un momento
determinado de un RT y, por consiguiente, su calidad biológica.
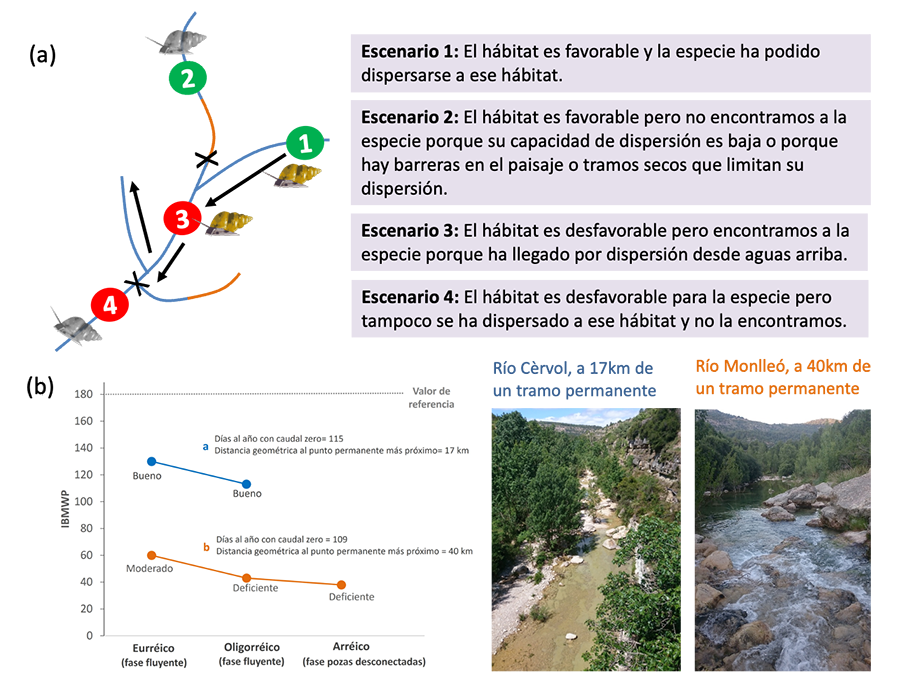
Figura 5. (a) Escenarios posibles para el
desarrollo de índices biológicos en ríos temporales teniendo en cuenta que las
comunidades se explican por una combinación de factores relacionados con el
nicho ecológico y la dispersión. Tradicionalmente, se han asumido los escenarios
1 y 4 en la aplicación de los índices biológicos, pero los escenarios 2 y 3
podrían influenciar el rendimiento de estos si no se considera la aproximación
de metacomunidades, ya sea por infravaloración (escenario 2) o sobrevaloración
(escenario 3) de la calidad biológica. Los tramos en azul de la red fluvial
corresponden a tramos con agua (fluyentes o pozas) mientras que los marrones a
cauces secos. Los círculos verdes indican hábitats favorables para la especie
(en este caso un macroinvertebrado de la familia Hydrobiidae) mientras que los
círculos rojos son hábitats desfavorables. Las flechas indican posibles vías de
dispersión de la especie y las X barreras a esta dispersión (p.e. por falta de
agua o por cambios en las condiciones locales). La especie se presenta en color
cuando está presente y en escala de grises cuando está ausente. (b)
Efecto sobre los índices biológicos (IBMWP) en dos ríos temporales, uno
relativamente conectado a un tramo permanente (Río Cèrvol, cuenca del Cèrvol) y
otro muy aislado (Río Monlleó, cuenca del Mijares). Los valores del índice se
presentan para la fase fluyente (estado acuático eurréico y oligorréico) y la
fase de pozas desconectadas (estado acuático arréico) con relación al valor de
referencia según su tipología y las respectivas clases de calidad. Adaptado de Cid et al.
(2020).
Figure
5. (a) Possible scenarios for the development of
biological indices in temporary rivers considering that communities are
explained by a combination of factors related to ecological niche and
dispersal. Traditionally, scenarios 1 and 4 have been assumed in the application
of biological indices, but scenarios 2 and 3 could influence their performance
if the metacommunity approach is not considered, either through undervaluation
(scenario 2) or overvaluation (scenario 3) of biological quality. The blue sections
of the river network correspond to segments with water (flowing or pools),
while the brown sections represent dry riverbeds. Green circles indicate
favorable habitats for the species (in this case, a macroinvertebrate of the
Hydrobiidae family), while red circles represent unfavorable habitats. Arrows
indicate possible dispersal routes of the species, and X marks represent
barriers to this dispersal (e.g., due to lack of water or changes in local
conditions). The species is shown in color when present and in grayscale when
absent. (b) Effect on
biological indices (IBMWP) in two temporary rivers, one relatively connected to
a permanent reach (Cèrvol river, Cèrvol basin) and another highly isolated
(Monlleó River, Mijares basin). Index values are presented for the flowing
phase (eurheic and oligorheic aquatic states) and the disconnected pools phase
(arheic aquatic state) in relation to the reference value according to their
typology and respective quality classes. Adapted from Cid et al. (2020).
La forma en que se
debe incorporar la combinación de factores locales y regionales en la
determinación del estado ecológico solo se ha abordado teóricamente (Siqueira et al. 2014; Cid et
al. 2020, 2021), pero su aplicación práctica pasa
por considerar los factores regionales en sentido amplio, tanto la capacidad de
dispersión de los organismos (espacial y temporal; Bonada
et al. 2017) como el grado de conectividad de cada sitio de muestreo (p.e.
aislamiento topográfico o conectividad hidrológica espaciotemporal (Cañedo-Argüelles et al. 2015; Cunillera-Montcusí et al. 2023). Por
ejemplo, para macroinvertebrados existe información sobre la capacidad de
dispersión espacial (Sarremejane et al. 2020)
y temporal (es decir, resistencia a la temporalidad del flujo; Bonada et al. 2017) que, juntamente con variables del
grado de conectividad paisaje e hidrológica, podrían utilizarse para
desarrollar un modelo predictivo tipo RIVPACS (Poquet
et al. 2009) a partir de unas comunidades de referencia o mínimamente
impactadas (Cid et al. 2020). Este modelo podría
predecir cómo de diferentes son las comunidades observadas (O) de un punto de
muestreo respecto a las esperadas en condiciones de referencia (E) según los
factores locales y regionales, y evaluar el estado ecológico a partir de la ratio
entre O/E. Valores de O/E más cercanos a 1 indicarían que O y E son similares y
que las comunidades del punto de muestreo que se evalúa están cerca de las
condiciones de referencia. En cambio, valores de O/E más cercanos a cero
indicarían que las comunidades sujetas a impactos humanos independientemente de
los factores regionales. Esta metodología se podría aplicar para cualquier
índice o métrica, como por ejemplo el índice IBMWP, calculando el valor de este
índice con las comunidades de referencia (E) y luego estableciendo clases de
calidad respecto a E y a lo largo de un gradiente de impacto antropogénico (Cid et al. 2020). Alternativamente, otra aproximación a
testar para incorporar de manera simultánea los factores locales con los
regionales, sería asignar un valor de capacidad de dispersión a cada familia,
tal y como el IBMWP asigna un valor de tolerancia a la contaminación, y combinando
los dos tipos de puntuaciones por familia.
Oportunidades para la evaluación del estado ecológico en
ríos temporales
La aprobación de la DMA y su posterior implementación supuso
un avance en la investigación para desarrollar o actualizar metodologías de
evaluación del estado ecológico. Actualmente, por ejemplo, conocemos bien cómo
responden los índices biológicos en ríos permanentes, cómo cambian las
comunidades entre tipologías, sus condiciones de referencia, los cortes de las
clases de calidad, y tenemos metodologías para evaluar el estado
hidrogeomorfológico y fisicoquímico contrastadas (Birk et
al. 2012; Reyjol et al. 2014; Ollero et al. 2021). Sin embargo, estas metodologías
siguen actualizándose en función de los avances científicos del momento que,
básicamente, pasan por la consideración de herramientas moleculares que cada
vez están más al alcance de científicos y gestores, el uso de grandes bases de
datos existentes que permiten modelizaciones complejas, el desarrollo de
índices alternativos que informan sobre los efectos de las perturbaciones a
niveles de organización más allá del taxón, y la implicación de la sociedad en
la gestión, que aporta una perspectiva socioecológica en el proceso de
evaluación del estado ecológico. Todos estos aspectos ofrecen también
oportunidades a explorar en RTs con la finalidad de obtener metodologías
fiables que mejoren las debilidades de las existentes.
Nuevas herramientas al alcance: el uso del metabarcoding
En sistemas tan sensibles y
dinámicos como los RTs, las técnicas tradicionales de evaluación del estado
ecológico pueden llegar a ser invasivas en determinadas fases del río. Por
ejemplo, las pozas desconectadas pueden tener una diversidad reducida pero
clave para la recolonización después de la fase seca, con lo que el muestreo
biológico puede conllevar la perturbación significativa del hábitat y la
eliminación de una gran parte de la comunidad biológica. El uso de técnicas
moleculares no invasivas, tales como las herramientas de secuenciación masiva (metabarcoding),
han supuesto una auténtica revolución debido a su alta eficacia para obtener
la composición de especies de una comunidad sin necesidad de invertir esfuerzos
en la separación e identificación morfológica y, en el caso del DNA ambiental,
incluso detectar presencia de especies acuáticas en agua o sedimento sin tener
que tomar una muestra biológica (Hering et al. 2018).
Estas técnicas, además, permiten no solo obtener
información a un nivel taxonómico menor (género o especie), sino también
identificar taxones alternativos que pudieran usarse como bioindicadores de
forma rutinaria (p.e. la meiofauna bentónica; Alves
et al. 2013). Con el tiempo, se espera también
que el uso de las herramientas moleculares permita desarrollar índices basados
en la expresión de genes implicados en los mecanismos para tolerar
perturbaciones (p.e. en la osmoregulación para evaluar los efectos de la
salinización; Hering et al. 2018), lo que podría ser muy interesante en RTs para
distinguir los efectos de la temporalidad de las perturbaciones antrópicas (Blackman et al. 2021).
A pesar de que el uso del metabarcoding
cada vez está más al alcance tanto a nivel técnico como económico, todavía
falta información de base para poderse aplicar de manera rutinaria por las
administraciones, y se requerirán esfuerzos significativos para la
estandarización y armonización de metodologías (Hering
et al. 2018). Además, el uso de estas técnicas
aún presenta dos limitaciones. Por un lado, las estimaciones de las abundancias
son todavía imprecisas, lo que limita la aplicación de índices cuantitativos,
indispensables en la implementación de la DMA. Por otro lado, si bien es cierto
que las librerías a niveles taxonómicos superiores están bastante completas,
existe todavía mucha falta de información cuando se pretenden asignar
secuencias de DNA a una especie concreta. Para la península ibérica, por
ejemplo, las librerías para macroinvertebrados solamente consideran un 35% de
las especies, pero un 93% de las familias (Múrria et
al. 2020). Ello no sería muy problemático para su uso en los índices
estandarizados que usan el nivel de familia (p.e. el IBMWP), pero sí si se
pretenden desarrollar índices más adecuados basados en géneros o especies, no
solamente para los RTs. A pesar de ello, también
existen limitaciones relacionadas con la estimación de la abundancia o el
origen del DNA muestreado (Deiner et al. 2017), e incertidumbres sobre si el metabarcoding debe
hacerse de la muestra global (bulk sample) o puede hacerse de otro tipo
de muestras (p.e. deriva, agua, eDNA) para
reducir los tiempos de recolección y procesado. La situación es más
dramática con las diatomeas, para las que las librerías solamente incluyen
información para un número muy reducido de especies debido a su carácter
unicelular y a las dificultades de aislamiento y
cultivo que presentan (Hering et al. 2018). En este caso el uso de OTUs (Operational Taxonomic
Units) o, preferiblemente, ESVs (Exact Sequence Variants, Tapolczai et al. 2021)
sería posible y facilitaría el desarrollo de índices más específicos (Pérez-Burillo et al. 2021), que requerirían sus correspondientes condiciones de referencia (Hering et al. 2018). Para
peces, el DNA ambiental ha resultado ser muy útil para detectar la presencia de
especies, y se presenta como una buena alternativa a las metodologías
estandarizadas (Hering et al. 2018). En la península ibérica, por ejemplo, las
librerías incluyen un 98,11% de las especies de peces de agua dulce presentes (Múrria et al. 2020).
Índices
alternativos para la evaluación del estado ecológico
A pesar de que la DMA contempla en su definición de estado
ecológico tanto la parte estructural como funcional de los ecosistemas, la
evaluación del estado ecológico se ha llevado a cabo teniendo en cuenta la
parte estructural, es decir, la información taxonómica de los organismos ya sea
a nivel de especie, género o familia. Si bien esta es la información básica
para poder realizar una evaluación mínima de un tramo fluvial, dicha
información puede ser complementada con información sobre los rasgos biológicos
de estos taxones, lo que permite evaluar la comunidad también desde el punto de
vista funcional, así como cuantificar la capacidad de resistencia y resiliencia
de las comunidades frente a perturbaciones (naturales o antrópicas). Además, la
consideración de múltiples rasgos biológicos permite entender cómo afectan los
diferentes tipos de perturbaciones a los organismos (p.e. el tamaño corporal se
relaciona con la contaminación por metales, mientras que la forma de movimiento
puede hacerlo con una perturbación hidrológica; Statzner
y Bêche 2010), mientras que los índices actuales se desarrollaron para
responder a la contaminación orgánica (Clarke et al.
2011). En la actualidad existen varios índices basados en rasgos
funcionales (p.e. Petchey y Gaston 2006; Legras et al. 2018; Soria et
al. 2020) que podrían ser incorporados a los protocolos actuales de
evaluación del estado ecológico, también para RTs. De hecho, la redundancia
funcional (basada en rasgos biológicos) de toda la comunidad de
macroinvertebrados o la presencia-ausencia de filtradores han mostrado detectar
perturbaciones antrópicas tanto en ríos permanentes como en RTs (Soria et al. 2020). También en macrófitos, el uso de
los rasgos biológicos y ecológicos está dando buenos resultados para posibles
aplicaciones en biomonitoring (Stefanidis et
al. 2023). Sin embargo, el uso generalizado y rutinario de estos índices
funcionales por parte de la administración pasa por (1) establecer protocolos
sencillos para poder utilizarlos (a menudo más complejos de obtener), (2)
disponer de bases de datos completas para una amplia batería de rasgos
funcionales que respondan a diferentes perturbaciones (naturales o antrópicas),
y (3) obtener información completa a nivel de género o de especie para cada uno
de los rasgos. En los últimos años ha habido intentos de generar bases de datos
de rasgos biológicos para diferentes grupos taxonómicos (p.e. Jeliazkov et al. 2020), las cuales serán muy útiles
para este fin, pero aún están muy incompletas y no incluyen rasgos que,
potencialmente, podrían responder a perturbaciones específicas.
Más allá del uso de rasgos funcionales, existen
aproximaciones que informan sobre los efectos de las perturbaciones a otros
niveles de organización, que podrían explorarse en RTs y para las que también
se ha sugerido su consideración en la DMA (Milinkovitch
et al. 2019). Los índices basados en biomarcadores moleculares, bioensayos
o la asimetría fluctuante, por ejemplo, nos informan sobre posibles efectos
subletales, y podrían responder a las perturbaciones antrópicas
independientemente de la temporalidad. Asimismo, estos índices podrían ser
complementarios a los rutinarios, tal y como se ha mostrado para ríos
permanentes con las especies diana Daphnia magna o Hydropsyche
exocellata (Damasio et al. 2011), y también
podrían ofrecer información relevante para los RTs.
Otras metodologías, como los índices basados en variables
sobre funcionamiento (p.e. descomposición de hojarasca) nos informan acerca de
los efectos a nivel ecosistémico, y son muy buenos candidatos para la
evaluación de efectos de las perturbaciones antrópicas (Bonada
et al. 2006). Sin embargo, estas variables sobre funcionamiento son, a
menudo, sensibles a la temporalidad o a sus consecuencias (p.e. la
descomposición de hojarasca incrementa con los nutrientes, pero también con la
temperatura, que suele ser más alta en RTs; Ferreira
et al. 2015), con lo que su uso rutinario en RTs para la evaluación del
estado ecológico es todavía incipiente y requeriría de calibración o adaptaciones
complejas.
Bases de datos
masivas producto de colaboración internacional y sensores remotos
En los últimos años han proliferado las colaboraciones
científicas internacionales destinadas a la recopilación de grandes bases de
datos (fruto de décadas de investigación) que cubren amplios gradientes
temporales y espaciales. Muchas de estas bases de datos incluyen RTs y podrían
ser utilizadas para testar hipótesis ecológicas y desarrollar estrategias
específicas para su gestión y conservación. Por ejemplo, existen modelos
hidrológicos que informan acerca de las redes hidrográficas mundiales con gran
precisión (Amatulli et al. 2022), así como
modelos que predicen la distribución de RTs a nivel global (Messager et al. 2021)
o el uso de datos de satélite para predecir el régimen hidrológico de los RTs (Cavallo et al. 2022; De
Girolamo et al. 2022). Al mismo tiempo, disponemos de datos acerca de la
abundancia y distribución global de insectos acuáticos (Grigoropoulou et al. 2023), peces (Comte et al. 2021) y otros muchos organismos asociados
a los RTs (Dornelas et al. 2018) a escala global
y, en muchos casos, durante largos períodos de tiempo. Aunque empieza a haber
trabajos que utilizan estas grandes bases de datos para abordar la ecología y
gestión de los ecosistemas fluviales (Jupke et al. 2022),
no existen aún estudios específicos para RTs más allá de algunas comparativas
intercontinentales (Bonada et al. 2008; Datry et al. 2014; Vander
Vorste et al. 2021). Cabe destacar también la existencia de proyectos de
investigación internacionales enfocados en RTs (Datry et
al. 2021) que abarcan amplios gradientes ambientales y que generarán datos
claves para mejorar nuestra comprensión y manejo de estos ecosistemas. Todo
ello se suma a la facilidad actual en el uso de herramientas de modelización
complejas, por ejemplo, técnicas de aprendizaje automático (del inglés “machine
learning”) que están siendo aplicadas en el desarrollo de metodologías para
la evaluación de la calidad biológica (Keck et al. 2017).
Ciencia ciudadana: implicación social en la
evaluación del estado hidrológico y ecológico
Puesto que la mayoría de RTs no forman parte de las redes
oficiales de monitoreo, la integración del mayor número posible de fuentes de
información es clave para mejorar la evaluación del estado hidrológico y
ecológico de estos ecosistemas. En este sentido, la ciencia ciudadana es un
buen recurso para generar conocimiento científico complementario y fiable para
los gestores (When et al. 2015; Krabbenhoft y Kashian 2020). Actualmente,
existen algunos ejemplos de herramientas de ciencia ciudadana que proporcionan
datos relacionados con la hidrología y la ecología de los RTs y que pueden
ayudar en su evaluación y gestión (Tabla 2).
Más allá de generar
conocimiento científico, la ciencia ciudadana también juega un papel esencial
fomentando la adquisición de nuevos conocimientos y habilidades por parte de la
ciudadanía (alfabetización científica y ambiental) (Turrini
et al. 2018). Además, crea una mayor conciencia sobre la biodiversidad y el
valor ecológico de los ecosistemas (Soria et al. 2021)
y, consecuentemente, facilita la participación ciudadana en procesos de toma de
decisiones relevantes en política ambiental (Turrini et
al. 2018). Esta capacidad educativa y transformadora de la ciencia
ciudadana puede ser clave en el caso de los RTs, ya que, en general, existe una
percepción social negativa en torno a los RTs, y estos son generalmente
considerados menos valiosos que los ríos permanentes (Rodríguez-Lozano et al. 2020). A esta
percepción social negativa de los RTs debemos añadir la evidencia científica de
que una visión simplificada de los ecosistemas fluviales suele estar
relacionada con una preferencia de medidas de gestión contrarias a la
sostenibilidad ambiental (Ladrera et al. 2020).
Para que la gestión y conservación de los RTs sea realmente efectiva, no sólo
se precisa mejorar las herramientas para su evaluación, sino que también se
necesita una participación activa e informada por parte de la sociedad que
impulse cambios en la legislación. En este contexto, la ciencia ciudadana
parece ser un mecanismo útil para promover la participación en la gestión de
los ecosistemas fluviales (When et al. 2015; Krabbenhoft y Kashian
2020), incluyendo los RTs (Soria et al. 2021).
Tabla 2. Herramientas
de ciencia ciudadana que consideran aspectos hidrológicos y/o ecológicos de los
ríos temporales.
Table
2. Citizen science tools that consider hydrological
and/or ecological aspects of temporary rivers.
La evaluación del
estado ecológico, de la teoría a la práctica
En este trabajo se han identificado 7 retos a tener en
cuenta para la correcta evaluación del estado ecológico de los RTs y 5
oportunidades que podrían contribuir, específicamente, a hacer frente a estos
retos (Fig. 6). En cualquier caso, la
conservación y correcta gestión de los RTs pasa por la evaluación objetiva y
fiable de su estado ecológico. Este proceso requiere abordar los retos
anteriormente indicados a nivel científico y el desarrollo de herramientas de
complejidad asumible para que su uso generalizado sea viable. Ello, de por sí
complejo en ríos permanentes, se revela todavía más difícil en RTs por su
propia naturaleza heterogénea y altamente cambiante, tanto a nivel espacial
como temporal. Así, por ejemplo, si bien la diversidad tipológica existente en
España ya es elevada (37 tipos actualmente definidos), al introducir el
elemento hidrológico (sintetizado en 4 hidrotipos: ríos permanentes, ríos
temporales fluyentes, ríos temporales estancados y ríos ocasionales o
episódicos), el grado de complejidad incrementa. Esta situación apunta a que es
necesario llevar a cabo una simplificación que permita abordar esta complejidad
de forma práctica. Así mismo, a pesar de que la obtención de índices de
evaluación del estado ecológico para RTs se augura compleja por todo lo
anteriormente expuesto, las metodologías que acaben derivando de las
investigaciones científicas actuales deberían considerar explícitamente que el
éxito de su implementación por parte de los gestores, y de su aceptación por
parte de la ciudadanía, se basa en su simplicidad, así como en la relación
coste-eficiencia. Esta implementación es fundamental teniendo en cuenta que las
predicciones de cambio global sugieren un aumento en la extensión y duración de
los RTs (Döll y Schmied 2012), también fuera de
la región de clima mediterráneo, con lo que el desarrollo y uso de nuevas
metodologías adaptadas a RTs constituye uno de los principales retos a asumir
en los próximos años.
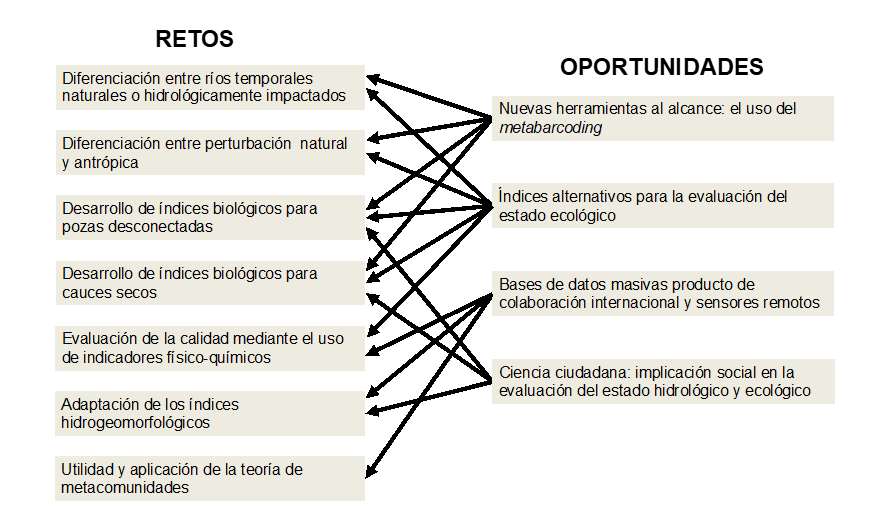
Figura 6.
Retos identificados para la para la evaluación del estado ecológico en ríos
temporales y como las diferentes oportunidades podrían contribuir a cada uno de
los retos.
Figure 6. Challenges identified for the evaluation of the ecological status
in temporary rivers and how the different opportunities could contribute to
each of the challenges.
Agradecimientos
Este estudio está financiado por el proyecto “DRY-Guadalmed:
Herramientas avanzadas para la evaluación del estado ecológico de ríos
temporales mediterráneos durante la fase seca” (PID2021-126143OB-C21 y
PID2021-126143OB-C22) financiado por el Ministerio de Ciencia e Innovación
(MCIN/AEI/10.13039/501100011033/) y por FEDER Una manera de hacer Europa.
Contribución de los autores
Núria Bonada:
Conceptualización, Adquisición de fondos, Administración del proyecto,
Supervisión, Redacción. Núria Cid: Conceptualización, Adquisición de fondos, Supervisión,
Redacción. María del Mar Sánchez Montoya: Conceptualización, Adquisición de
fondos, Administración del proyecto, Supervisión, Redacción. Raúl Acosta:
Redacción. María Isabel Arce: Redacción. Miguel Cañedo-Argüelles: Redacción.
David Cunillera-Montcusí: Redacción. Paloma de las Heras: Redacción. María
Díaz-Redondo: Redacción. Zeynep Ersoy: Redacción. José María Fernández-Calero:
Redacción. Núria Flor Arnau: Redacción. Pau Fortuño: Redacción. Zeus Freixinos:
Redacción. Joan Gomá: Redacción. Antonio López-Pintor: Redacción. Manuel Jesús
López-Rodríguez: Redacción. Nieves López-Rodríguez: Redacción. Andrés
Mellado-Díaz: Redacción. Jesús Miñano-Martínez: Redacción. Felipe Morcillo:
Redacción. Antoni Munné: Redacción. Marc Ordeix: Redacción. Martí Piñero:
Redacción. Guillermo Quevedo-Ortiz: Redacción. Neftalí Roblas: Redacción. Pablo Rodríguez-Lozano: Redacción. José Vicente Rovira: Redacción. Carlota Sánchez-Campaña: Redacción. Judit Sánchez-Nogueras: Redacción. Maria Soria: Redacción. Mª Luisa Suárez Alonso: Redacción.
José Manuel Tierno de Figueroa: Redacción.
Maria Rosario Vidal-Abarca: Redacción.
Dolors Vinyoles: Redacción. Francesc
Gallart: Redacción. Narcís Prat: Redacción.
Referencias
Acuña, V., Datry, T., Marshall, J., Barceló, D., Dahm, C.N.,
Ginebreda, A., McGregor, G., et al. 2014. Why should we care about temporary waterways? Science
343(6175): 1081-1081.
Alves, A.S., Adão, H., Ferrero, T.J., Marques,
J.C., Costa, M.J., Patrício, J. 2013. Benthic meiofauna as indicator of
ecological changes in estuarine ecosystems: The use of nematodes in ecological
quality assessment. Ecological Indicators 24: 462-475.
Amatulli, G., Garcia Marquez, J., Sethi, T., Kiesel, J., Grigoropoulou, A.,
Üblacker, M.M., Shen, Q.S., et al. 2022. Hydrography90m: A new high-resolution
global hydrographic dataset. Earth System Science Data 14(10):
4525-4550.
Aparicio, E., Carmona‐Catot,
G., Moyle, P.B., García‐Berthou, E. 2011. Development and evaluation of a fish‐based index to assess biological integrity
of Mediterranean streams. Aquatic Conservation: Marine and Freshwater
Ecosystems 21(4): 324-337.
Arce, M.I., Sánchez-Montoya, M.M., Vidal-Abarca,
M.R., Suárez, M.L., Gómez, R. 2014. Implications of flow intermittency on
sediment nitrogen availability and processing rates in a Mediterranean
headwater stream. Aquatic Sciences 76: 173-186.
Arce, M.I., Mendoza-Lera, C., Almagro, M., Catalán, N.,
Romaní, A.M., Martí, E., Gómez, R., et al. 2019. A conceptual framework for understanding the
biogeochemistry of dry riverbeds through the lens of soil science. Earth-Science
Reviews 188: 441-453.
Arce, M.I., Sánchez-García, M., Martínez-López,
J., Cayuela, M.L., Sánchez-Monedero, M.A. 2023. Role of dry watercourses of an
arid watershed in carbon and nitrogen processing along an agricultural impact
gradient. Journal of Environmental Management 333: 117462.
Barton, P.S., Westgate, M.J., Foster, C.N.,
Cuddington, K., Hastings, A., O’Loughlin, L.S., Sato, C.F., et al. 2020. Using
ecological niche theory to avoid uninformative biodiversity surrogates. Ecological
Indicators 108: 105692.
Bailey, R.C., Norris, R.H., Reynoldson, T.B.
(eds.) 2004. Bioassessment of Freshwater Ecosystems: Using the Reference
Condition Approach. Springer, Nueva
York, NY, EE.UU.
Belmar, O., Bruno, D.,
Guareschi, S., Mellado‐Díaz, A., Millán, A., Velasco, J. 2019. Functional
responses of aquatic macroinvertebrates to flow regulation are shaped by
natural flow intermittence in Mediterranean streams. Freshwater Biology
64(5): 1064-1077.
Benejam, L., Saura-Mas, S., Bardina, M., Solà, C.,
Munné, A., García-Berthou, E. 2016. Ecological impacts of small hydropower
stations on headwater stream fish: from individual to community effects. Ecology
of Freshwater Fish 25(2): 295-306.
Birk, S., Bonne, W., Borja, A., Brucet, S.,
Courrat, A., Poikane, S., Solimini, A., et al. 2012. Three hundred ways to
assess Europe's surface waters: an almost complete overview of biological
methods to implement the Water Framework Directive. Ecological Indicators
18: 31e41.
Blackman, R.C., Altermatt, F., Foulquier, A.,
Lefébure, T., Gauthier, M., Bouchez, A., Stubbington, R., et al. 2021.
Unlocking our understanding of intermittent rivers and ephemeral streams with
genomic tools. Frontiers of Ecology and the Environment 19: 574-583.
Bogan, M.T., Chester, E.T., Datry, T., Murphy,
A.L., Robson, B.J., Ruhí, A., Stubbington, R., et al. 2017. Resistance,
resilience, and community recovery in intermittent rivers and ephemeral
streams. En: Datry, T., Bonada N., Boulton, A.J. (eds.) Intermittent Rivers
and Ephemeral Streams: Ecology and Management, pp. 349-376. Academic Press, Londres, Reino Unido.
Bonada, N., Resh, V.H. 2013. Mediterranean-climate
streams and rivers: geographically separated but ecologically comparable
freshwater system. Hydrobiologia 719: 1-29.
Bonada, N., Prat, N., Resh, V.H., Statzner, B.
2006. Developments in aquatic insect biomonitoring: a comparative analysis of
recent approaches. Annual Review of Entomology 51: 495-523.
Bonada, N., Rieradevall, M., Prat, N. 2007.
Macroinvertebrate community structure and biological traits related to flow
permanence in a Mediterranean river network. Hydrobiologia 589: 91-106.
Bonada, N., Rieradevall, M., Dallas, H., Davies, J., Day, J.,
Figueroa, R., Resh, V.H., et al. 2008. Multi-scale assessment of macroinvertebrate richness
and composition in Mediterranean-climate rivers. Freshwater Biology 53:
772–788.
Bonada, N., Carlson, S.M., Datry, T., Finn, D.S.,
Leigh, C., Lytle, D.A., Monaghan, M.T., et al. 2017. Genetic, evolutionary, and
biogeographical processes in intermittent rivers and ephemeral streams. En:
Datry, T.; Bonada, N., Boulton, A.J. (eds.), Intermittent Rivers and
Ephemeral Streams: Ecology and Management, pp. 405-431. Academic Press,
Londres, Reino Unido.
Bonada, N., Cañedo-Argüelles, M., Gallart, F., Von
Schiller, D., Fortuño, P., Latron, J., Llorens, P., et al. 2020. Conservation
and management of isolated pools in temporary rivers. Water 12: 2870.
Brown, B.L.,
Swan, C.M. 2010. Dendritic
network structure constrains metacommunity properties in riverine ecosystems. Journal
of Animal Ecology 79: 571–580.
Cañedo‐Argüelles, M., Boersma, K. S., Bogan, M.T., Olden, J.D.,
Phillipsen, I., Schriever, T.A., Lytle, D.A. 2015. Dispersal strength
determines meta‐community structure in a dendritic riverine
network. Journal of Biogeography 42(4): 778-790.
Cavallo, C., Papa, M.N., Negro, G., Gargiulo, M.,
Ruello, G., Vezza, P. 2022. Exploiting Sentinel-2 dataset to assess flow
intermittency in non-perennial rivers. Scientific Reports 12: 21756.
Cid, N., Verkaik, I., García-Roger, E.M.,
Rieradevall, M., Bonada, N., Sánchez-Montoya, M.M., Gómez, R., et al. 2016. A
biological tool to assess flow connectivity in reference temporary streams from
the mediterranean basin. Science of the Total Environment 540: 178-190.
Cid, N., Bonada, N., Heino, J.,
Cañedo-Argüelles, M., Crabot, J., Sarremejane, R., Soininen, J., et al. 2020. A
metacommunity approach to improve biological assessments in highly dynamic
freshwater ecosystems. BioScience 70(5): 427-438.
Cid, N., Erös, T., Heino, J., Singer, G., Jähnig, S.C.,
Cañedo-Argüelles, M., Bonada, N., et al. 2021. From meta-system theory to the sustainable management
of rivers in the Anthropocene. Frontiers in Ecology and the Environment
20: 49-57.
Clarke, R.T., Davy-Bowker, J., Dunbar, M., Laize,
C., Scarlett, P., Murphy, J. 2011. SNIFFER
WFD119: Enhancement of the River Invertebrate Classification Tool (RICT). Project Report. Scotland & Northern Ireland
Forum for Environmental Research. Edinburgh, Scotland. Reino Unido.
Chiu, M.C., Leigh, C., Mazor, R., Cid, N., Resh, V.H. 2017.
Anthropogenic threats to
intermittent rivers and ephemeral streams. En: Datry, T., Bonada, N., Boulton,
A.J. (eds.), Intermittent Rivers and Ephemeral Streams: Ecology and
Management, pp. 433-454. Academic Press, London, Reino Unido.
Comisión Europea. 2000. Directive 2000/60/EC of October 23,
2000, of the European Parliament and of the Council establishing a framework
for community action in the field of water policy. Official Journal of the
European Union L327: 1-72.
Comisión
Europea. 2011. Guidance
document on the intercalibration process 2008-2011. Guidance document No 14, European Commission. Disponible en: https://data.europa.eu/doi/10.2779/99432
Comte, L.,
Carvajal‐Quintero, J., Tedesco, P. A., Giam, X., Brose, U.,
Erős, T., Filipe, A.F., et al. 2021. RivFishTIME: A global database of fish time‐series to study global change ecology in
riverine systems. Global Ecology and Biogeography 30(1): 38-50.
Corti, R.,
Datry, T. 2016. Terrestrial
and aquatic invertebrates in the riverbed of an intermittent river: Parallels
and contrasts in community organisation. Freshwater
Biology 61: 1308-1320.
Cunillera‐Montcusí, D.,
Fernández‐Calero, J.M., Pölsterl, S., Argelich, R., Fortuño, P.,
Cid, N., Bonada, N., et al. 2023. Navigating through space and time: A methodological approach to quantify
spatiotemporal connectivity using stream flow data as a case study. Methods
in Ecology and Evolution 14(7): 1780-1795.
Damásio, J., Fernández-Sanjuan, M., Sánchez-Avila,
J., Lacorte, S., Prat, N., Rieradevall, M., Soares, A.M., et al. 2011.
Multi-biochemical responses of benthic macroinvertebrate species as a
complementary tool to diagnose the cause of community impairment in polluted
rivers. Water Research 45: 3599-3613.
Datry, T., Larned, S.T., Fritz, K.M., Bogan,
M.T., Wood, P.J., Meyer, E.I., Santos, A.N. 2014. Broad-scale patterns of
invertebrate richness and community composition in temporary rivers: effects of
flow intermittence. Ecography 37: 94-104.
Datry, T., Boulton, A.J., Bonada, N., Fritz, K.,
Leigh, C., Sauquet, E., Tockner, K., et al. 2018. Flow intermittence and
ecosystem services in rivers of the Anthropocene. Journal of Applied Ecology
55: 353-364.
Datry, T., Allen, D., Argelich, R., Barquin, J.,
Bonada, N., Boulton, A., Branger, F., et
al. 2021. Securing biodiversity, functional integrity, and ecosystem services
in drying river networks (DRYvER). Research Ideas and Outcomes 7:
e77750.
Deiner, K., Bik, H.M., Mächler, E., Seymour, M., Lacoursière‐Roussel,
A., Altermatt, F., Creer, S.,
et al. 2017. Environmental DNA metabarcoding: transforming how we survey animal
and plant communities. Molecular
Ecology 26: 5872–95.
De Girolamo, A. M., Drouiche, A., Ricci, G. F., Parete, G.,
Gentile, F., Debieche, T. H. 2022. Characterising flow regimes in a semi-arid region with limited data
availability: the Nil Wadi case study (Algeria). Journal of Hydrology:
Regional Studies 41: 101062.
Delgado, C., Pardo, I., García, L. 2012. Diatom
communities as indicators of ecological status in Mediterranean temporary
streams (Balearic Islands, Spain). Ecological Indicators 15(1): 131-139.
Döll, P.,
Schmied, H.M. 2012. How is
the impact of climate change on river flow. Environmental Research Letters
7: 014037.
Dornelas, M., Antao, L.H., Moyes, F., Bates, A.E.,
Magurran, A.E., Adam, D., Akhmetzhanova, A.A.,
et al. 2018. BioTIME: A database of biodiversity time series for the
Anthropocene. Global Ecology and Biogeography 27(7): 760-786.
EFI+
CONSORTIUM 2009.
Manual for the application
of the new European Fish Index –EFI+. A fish-based method to assess the
ecological status of European running waters in support of the Water Framework
Directive. Disponible en: https://oa.upm.es/6346/
Feio, M.J., Ferreira, J., Buffagni, A., Erba, S.,
Dörflinger, G., Ferréol, M., Munné, A., et al. 2014. Comparability of ecological quality
boundaries in the Mediterranean basin using freshwater benthic invertebrates.
Statistical options and implications. Science of the Total Environment 476-477:
777-784.
Fellman, J.B., Dogramaci, S., Skrzypek, G., Dodson,
W., Grierson, P.F. 2011. Hydrologic control of dissolved organic matter
biogeochemistry in pools of a subtropical dryland river. Water Resources
Research 47: W06501.
Fernández, D., Barquin, J., Raven, P.J. 2011. A
review of river habitat characterisation methods: indices vs. characterisation
protocols. Limnetica 30(2): 217-234.
Ferreira, V., Castagneyrol, B., Koricheva, J.,
Gulis, V., Chauvet, E., Graça, M.A.S. 2015. A meta-analysis of the effects of
nutrient enrichment on litter decomposition in streams. Biological Reviews
90: 66-688.
Florsheim, J.L., Mount, J.F., Chin, A. 2008. Bank
erosion as a desirable attribute of rivers. Bioscience 58: 519-529.
Fritz, K.M., Glime, J.M., Hribljan, J.,
Greenwood, J.L. 2009. Can bryophytes be used to characterize hydrologic
permanence in forested headwater streams? Ecological Indicators 9:
681-692.
Fritz, K.M., Kashuba, R.O., Pond, G.J.,
Christensen, J.R., Alexander, C., Washington, B.J., Johnson, B.R., et al. 2023.
Identifying invertebrate indicators for streamflow duration assessments in
forested headwater streams. Freshwater Science 42: 247-267.
Gallart, F., Prat, N., García-Roger, E.M., Latron,
J., Rieradevall, M., Llorens, P., Barberá, G.G., et al. 2012. A novel approach
to analysing the regimes of temporary streams in relation to their controls on
the composition and structure of aquatic biota. Hydrology and Earth System
Sciences 16: 3165-3182.
Gallart, F., Cid, N., Latron, J., Llorens, P.,
Bonada, N., Jeuffroy, J., Jiménez-Argudo, S-M., et al. 2017. TREHS: An
open-access software tool for investigation and evaluating temporary river
regimes as a first step for their ecological status assessment. Science of the Total Environment 607-608: 519-540.
García, C.,
Rodríguez-Lozano, P. 2020. Cinco
aspectos clave en la gestión del agua de las Islas Baleares. En: Santamarta,
J.C., Rodríguez-Martín, J. (eds.), Los procesos de planificación hidrológica
en la península ibérica e islas en un contexto de cambio climático, pp.
231-246. Colegio Oficial de Ingenieros de Montes, Madrid, España.
García, L., Pardo, I., Delgado, C. 2014. Macroinvertebrate indicators of ecological
status in Mediterranean temporary stream types of the Balearic Islands. Ecological Indicators 45: 650-663.
García-Berthou, E., Bae, M., Benejam, L., Alcaraz, C., Casals, F., De
Sostoa, A., Solá, C., et al. 2016. Fish-based indices in Catalan rivers: Intercalibration and comparison of
approaches. En: Munné, A., Ginebreda, A., Prat,
N. (eds.), Experiences from Surface Water Quality Monitoring: The EU Water
Framework Directive Implementation in the Catalan River Basin District (Part I),
pp.125-147. Springer Verlag, Berlin,
Heidelberg, Alemania.
Gerlach, J., Samways, M., Pryke,
J. 2013. Terrestrial invertebrates as bioindicators: an overview of available
taxonomic groups. Journal of Insect Conservation 17: 831-850.
Gómez, R., Arce, M.I., Baldwin, D.S., Dahm, C.N.
2017. Water physicochemistry in intermittent rivers and ephemeral streams. En: Intermittent
Rivers and Ephemeral Streams: Ecology and Management, Datry, T., Bonada,
N., Boulton, A.J. (eds.), pp. 109-134. Academic Press, London, Reino Unido.
Grigoropoulou, A., Ab Hamid, S., Acosta, R., Akindele,
E.O., Al-Shami, S.A., Altermatt, F., Amatulli, G., et al. 2023. The global EPTO
database: Worldwide occurences of aquatic insects. Global Ecology and
Biogeography 32(5): 642-655.
Heino, J. 2013. Does dispersal ability affect the
relative importance of environmental control and spatial structuring of
littoral macroinvertebrate communities? Oecologia 171: 971-980.
Hering, D., Borka, A., Jones, J.I., Pont, D.,
Boets, P., Bouchez, A., Bruce, K., et al. 2018. Implementation options for
DNA-based identification into ecological status assessment under the European
Water Framework Directive. Water Research 138: 192-205.
Hermoso, V., Clavero, M., Blanco-Garrido, F., Prenda, J. 2010.
Assessing the ecological status
in species-poor systems: A fish-based index for Mediterranean rivers (Guadiana
River, SW Spain), Ecological Indicators 10(6): 1152–1161.
Hodkinson, I.D., Jackson, J.K. 2005. Terrestrial and aquatic
invertebrates as bioindicators for environmental monitoring, with particular
reference to mountain ecosystems. Environmental Management 35: 649-666.
Hubbell, S.P. 2001. The Unified Neutral Theory
of Biodiversity and Biogeography. Princeton University Press, Princeton, EEUU.
IPBES. 2019. Global Assessment Report on
Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy
Platform on Biodiversity and Ecosystem Services. Disponible en: https://www.ipbes.net/global-assessment
IPCC. 2022. Climate Change 2022: Impacts,
Adaptation and Vulnerability. The Working Group II contribution to the IPCC
Sixth Assessment Report assesses the impacts of climate change, looking at
ecosystems, biodiversity, and human communities at global and regional level.
Intergovernmental panel on climate change (IPCC). Disponible en: https://www.ipcc.ch/report/ar6/wg2/
Jeliazkov, A., Mijatovic, D., Chantepie, S., Andrew,
N., Arlettaz, R., Barbaro, L., Barsoum, N., et al. 2020. A global database for
metacommunity ecology, integrating species, traits, environment and space. Scientific Data 7(1):
1-15.
Jupke, J.F., Birk, S., Álvarez-Cabria, M., Aroviita, J.,
Barquín, J., Belmar, O., Bonada, N., et al. 2022. Evaluating the biological validity of European River
Typology Systems with least disturbed benthic macroinvertebrate communities. Science
of the Total Environment 842: 156689.
Keck, F., Vasselin, V., Tapolczai, K., Rimet,
F., Bouchez, A. 2017. Freshwater biomonitoring in the Information Age.
Frontiers in Ecology and the Environment 15: 266-274.
Krabbenhoft,
C.A., Kashian, D.R. 2020.
Citizen science data are a reliable complement to quantitative ecological
assessments in urban rivers. Ecological Indicators 116: 106476.
Labat, F., Thiébaut, G., Piscart, C. 2022. A new
method for monitoring macrophyte communities in small shallow lakes and ponds. Biodiversity Conservation 31: 1627-1645.
Ladrera, R., Rodríguez-Lozano, P., Verkaik, I., Prat, N.,
Díez, J.R. 2020. What do
students know about rivers and their management? Analysis by educational stages
and territories. Sustainability 12(20): 8719.
Lazaridou, M., Ntislidou, C., Karaouzas, I.,
Skoulikidis, N. 2016. Development of a national assessment method for the
ecological status of rivers in Greece, using the biological quality element
“benthic macroinvertebrates”, The Hellenic Evaluation System-2 (HESY-2), and
harmonisation with the results of the completed intercalibration of the Med GIG
(RM1, RM2, RM4, RM5), Ministry of Environment, Greece. p.30.
Legras, G., Loiseau, N., Gaertner, J.C. 2018. Functional richness: Overview of indices and
underlying concepts. Acta Oecologica 87: 34-44.
Leibold, M.A.,
Chase, J.M. 2018. Metacommunity
Ecology. Princeton University Press, Princeton, EEUU.
Lobera, G., Besné, P., Vericat, D., López-Tarazón, J.A.,
Tena, A., Aristi, I., Díez, J.R., et al. 2015. Geomorphic status of regulated rivers in the Iberian
Peninsula. Science of the Total Environment 508, 101-114.
Magalhães, M.F., Ramalho, C.E., Collares-Pereira,
M.J. 2008. Assessing biotic integrity in a Mediterranean watershed: Development
and evaluation of a fish-based index. Fisheries
Management and Ecology 15(4): 273-289.
MAGRAMA. 2015. Real Decreto 817/2015, de 11 de septiembre, por el
que se establecen los criterios de seguimiento y evaluación del estado de las
aguas superficiales y las normas de calidad ambiental. Ministerio de
Agricultura, Alimentación y Medio Ambiente. BOE, núm. 219, de 12 de
septiembre de 2015, págs.: 80582 a 80677.
Mandaric, L., Kalogianni, E., Skoulikidis, N.,
Petrovic, M., Sabater, S. 2019. Contamination patterns and attenuation of
pharmaceuticals. Science of the Total
Environment 647: 561-569.
MARM. 2008. Orden ARM/2656/2008, de 10 de septiembre, por la que
se aprueba la instrucción de planificación hidrológica. Ministerio de Medio
Ambiente, y Medio Rural y Marino. BOE núm. 229, de 22 de septiembre de
2008, págs.: 38472 a 38582.
Martín-Vide, J.P. 1999. Restoration of an urban river
in Barcelona, Spain. Environmental
Engineering and Policy 2(3): 113-119.
Martínez-Yoshino, N., Suárez-Alonso, M.L., Vidal-Abarca, M.R. 2021.
Delimitando los rasgos biológicos de la vegetación de los ríos secos: el caso
de las ramblas de la cuenca del Segura (SE de España). Anales de Biología
43: 11-25.
Mazor, R.D., Stein, E.D.,
Ode, P.R., Schiff, K. 2014. Integrating intermittent streams into watershed assessments:
Applicability of an index of biotic integrity. Freshwater Science 33(2):
459-474.
Mesnage, V., Bonneville, S., Laignel, B., Lefebvre,
D., Dupont, J.P., Mikes, D. 2002. Filling of a wetland (Seine estuary, France):
Natural eutrophication or anthropogenic process? A sedimentological and
geochemical study of wetland organic sediments. En: Nutrients and
Eutrophication in Estuaries and Coastal Waters: Proceedings of the 31st
Symposium of the Estuarine and Coastal Sciences Association (ECSA), held in
Bilbao, Spain, 3–7 July 2000, pp. 423-435). Springer, Netherlands.
Messager, M.L., Lehner, B., Cockburn, C., Lamouroux,
N., Pella, H., Snelder, T., Tockner, K., et al. 2021. Global prevalence of
non-perennial rivers and streams. Nature 594: 391-397.
Milinkovitch, T., Geffard, O., Geffard, A., Mouneyrac,
C., Chaumont, A., Xuereb, B., Fisson, C., et al. 2019. Biomarkers as tools for
monitoring within the Water Framework Directive context: concept, opinions and
advancement of expertise. Environmental
Science and Pollution Research 26:
32759-32763.
MITECO. 2019. Protocolo
de caracterización hidromorfológica de masas de agua de la categoría ríos,
código M-R-HMF-2019, 22 de abril de 2019. Ministerio para la Transición
Ecológica y el Reto Demográfico, anteriormente Ministerio para la Transición
Ecológica (MITECO), Madrid, España. Disponible en: https://www.miteco.gob.es/es/agua/temas/estado-y-calidad-de-las-aguas/aguas-superficiales/programas-seguimiento/protocolos-caracterizacion-y-calculo-metricas-en-hidromorfologia.html
MITECO. 2020. Guía para la evaluación del estado de las
aguas superficiales y subterráneas. Ministerio para la Transición Ecológica
y el Reto Demográfico. Madrid, España. .pp 403. Disponible en: https://www.miteco.gob.es/es/agua/temas/estado-y-calidad-de-las-aguas/medida-tomas-muestras.html
Moss, B. 2008. The Water Framework Directive: Total
environment or political compromise? Science of the Total Environment
400: 32–41.
Munné, A., Prat,
N. 2009. Use of
macroinvertebrate-based multimetric indices for water quality evaluation in
Spanish Mediterranean rivers: an intercalibration approach with the IBMWP
index. Hydrobiologia 628: 203-225.
Munné, A., Prat,
N. 2011. Effects of
Mediterranean-climate annual variability on stream biological quality
assessment using macroinvertebrate communities. Ecological Indicators 11: 651-662.
Munné, A., Prat, N., Solá, C., Bonada, N., Rieradevall, M.
2003. A simplified method to
assess ecological quality of riparian environment in rivers and streams: QBR
index. Aquatic Conservation: Marine and Freshwater Ecosystems 13:
147-163.
Munné, A., Bonada, N., Cid, N.,
Gallart, F., Solà, C., Bardina, M., Rovira, A., et al. 2021. A Proposal to
Classify and Assess Ecological Status in Mediterranean Temporary Rivers:
Research Insights to Solve Management Needs. Water 13: 767.
Múrria, C., Väisänen, L.O.S., Somma, S.,
Wangensteen, O.S, Arnedo, M.A., Prat, N. 2020. Towards an Iberian DNA barcode
reference library of freshwater macroinvertebrates and fishes. Limnetica 39:
73-92.
Ollero, A., Conesa, C, Vidal-Abarca, M.R. (eds.). 2021.
Buenas prácticas en gestión y restauración de cursos efímeros mediterráneos:
resiliencia y adaptación al cambio climático. Editum, Ediciones de la
Universidad de Murcia, España.
Papastergiadou,
E., Manolaki, P. 2012. Developing an Assessment System of RM-4
& RM-5 River Types for Cyprus Rivers. Final Report of Project ΤΑΥ 84/2009. Natural
Resources and Environment, Water Development Department, Ministry of
Agriculture, Cyprus.
Pérez-Burillo, J., Trobajo, R., Leira, M., Keck, F., Rimet, F.,
Sigró, J., Mann, D.G. 2021. DNA
metabarcoding reveals differences in distribution patterns and ecological
preferences among genetic variants within some key freshwater diatom species. Science
of the Total Environment 798: 149029.
Poff, N.L., Richter, B.D., Arthington, A.H.,
Bunn, S.E., Naiman, R.J., Kendy, E., Acreman, M.,
et al. 2010. The ecological limits of hydrologic alteration (ELOHA): a
new framework for developing regional environmental flow standards. Freshwater Biology 55: 147-170.
Poquet, J.M., Alba-Tercedor, J., Puntí, T., Sánchez-Montoya,
M.M., Robles, S., Álvarez, M., Zamora-Muñoz, C., et al. 2009. The MEDiterranean Prediction and
Classification System (MEDPACS): an implementation of the RIVPACS/AUSRIVAS
predictive approach for assessing Mediterranean aquatic macroinvertebrate
communities. Hydrobiologia 623, 153–171.
Petchey,
O.L., Gaston, K.J. 2006.
Functional diversity: Back to basics and looking forward. Ecology Letters
9(6): 741-758.
Reyjol, Y., Argillier, C., Bonne, W., Borja, A.,
Buijse, A.D., Cardoso, A.C., Daufresne, M., et al. 2014. Assessing the
ecological status in the context of the European Water Framework Directive:
Where do we go now? Science of the Total Environment 497-498: 332-344.
Rinaldi, M., Surian, N., Comiti, F., Bussettini, M.
2013. A method for the assessment and analysis of the hydromorphological
condition of Italian streams: The Morphological Quality Index (MQI). Geomorphology
180-181: 96-108.
Rodríguez-Lozano, P., Woelfle-Erskine, C., Bogan, M.T.,
Carlson, S.M. 2020. Are nonperennial rivers considered as valuable and worthy
of conservation as perennial rivers? Sustainability 12(14): 5782.
Rosindell, J., Hubbell, S.P., Etienne, R.S. 2011. The
Unified Neutral Theory of Biodiversity and Biogeography at Age Ten. Trends
in Ecology and Evolution 26(7): 340-348.
Sánchez-Montoya, M.M., von Schiller, D., Ruhí, D., Sting
Pechar, G., Proia, L., Miñano, J., Vidal-Abarca, M.R., et al. 2016. Responses
of ground-dwelling arthropods to surface flow drying in channels and adjacent
habitats along Mediterranean streams. Ecohydrology 9: 1376-1387.
Sánchez-Montoya, M.M., von Schiller, D., Barberá, G.G.,
Diaz, A.M., Arce, M.I., Del Campo, R., Tockner, K. 2018. Understanding the
effects of predictability, duration, and spatial pattern of drying on benthic
invertebrate assemblages in two contrasting intermittent streams. PLoS One
13(3): e0193933.
Sánchez-Montoya, M.M, Tockner,
K., von Schiller, D., Miñano, J., Catarineu, C., Lencina, J.L., Barberá, G.G.,
et al. 2020a. Dynamics of ground-dwelling arthropod metacommunities in
intermittent streams: The key role of dry riverbeds. Biological Conservation 241: 108328.
Sánchez-Montoya, M.M., Guerrero-Brotons, M., Miñano, J., Gómez, R.
2020b. Effects of debris piles
and pools along dry riverbeds on nutrients, microbial activity, and
ground-dwelling arthropods: A Namibian ephemeral river case. Journal of Arid
Environments 175: 104082.
Sánchez-Montoya, M.M., Gómez, R., Calvo, J.F., Bartonicka,
T., Datry, T., Paril, P. 2022. Ecological values of intermittent rivers for
terrestrial vertebrate fauna. Science of the Total Environment 806:
151308.
Sarremejane, R., Mykrä, H., Bonada, N., Aroviita, J., Muotka, T.
2017. Habitat connectivity and
dispersal ability drive the assembly mechanisms of macroinvertebrate
communities in river networks. Freshwater Biology 62(6): 1073-1082.
Sarremejane, R., Cid, N., Stubbington, R., Datry, T.,
Alp, M., Cañedo-Argüelles, M., Cordero-Rivera, A., et al. 2020. DISPERSE, a
trait database to assess the dispersal potential of European aquatic
macroinvertebrates. Scientific Data 7: 386.
Siqueira T., Durães L.D., de Oliveira Roque, F. 2014. Predictive modelling of insect
metacommunities in biomonitoring of aquatic networks. En: Ferreira, C.P, Godoy,
W.A.C. (eds.), Ecological Modelling Applied to Entomology, pp 109–126.
Springer International Publishing, Suiza.
Skoulikidis, N.T. 2008. Defining chemical status of a
temporary Mediterranean River. Journal of Environmental Monitoring
10(7): 842-852.
Soria, M., Leigh, C., Datry, T., Bini, L.M.,
Bonada, N. 2017. Biodiversity in perennial and intermittent rivers: a
meta-analysis. Oikos 126: 1078-1089.
Soria, M., Gutiérrez-Cánovas, C., Bonada, N., Acosta, R.,
Rodríguez-Lozano, P., Fortuño, P., Burgazzi, G., et al. 2020. Natural disturbances can produce misleading
bioassessment results: identifying metrics to detect anthropogenic impacts in
intermittent rivers. Journal of Applied Ecology 57: 283-295.
Soria, M., Bonada, N., Ballester, A., Verkaik,
I., Jordà-Capdevila, D., Solà, C., Munné, A.,
et al. 2021. Adapting participatory processes in temporary rivers management. Environmental
Science Policy 120, 145–156.
Statzner, B.,
Bêche, L.A. 2010. Can
biological invertebrate traits resolve effects of multiple stressors on running
water ecosystems? Freshwater Biology 55: 80-119.
Stefanidis, K., Dimitrellos, G., Sarika, M., Tsoukalas, D.,
Papastergiadou, E. 2022. Ecological quality assessment of Greek lowland
rivers with aquatic macrophytes in compliance with the EU Water Framework
Directive. Water 14: 2771.
Stefanidis, K., Oikonomou, A., Dimitrellos, G., Tsoukalas, D.,
Papastergiadou, E. 2023. Relationships
between environmental factors and functional traits of macrophyte assemblages
in running waters of Greece. Diversity 15: 949.
Steward, A.L., von Schiller, D.,
Tockner, K., Marshall, J.C., Bunn, S.E. 2012. When the rivers runs dry: Human
and ecological values of dry riverbeds. Frontiers in Ecology and the
Environment 10: 202–209.
Steward, A.L., Langhans, S.D., Corti, R., Datry, T.
2017. The biota of intermittent rivers and ephemeral streams: Terrestrial and
semi- aquatic invertebrates. En: Datry, T.; Bonada, N., Boulton, A.J. (eds). Intermittent
Rivers and Ephemeral Streams: Ecology and Management, pp 245-271. Academic
Press, Londres, Reino Unido.
Steward, A.L., Negus, P.,
Marshall, J.C., Clifford, S.E., Dent, C. 2018. Assessing the ecological health
of rivers when they are dry. Ecological Indicators 85: 537-547.
Steward, A.L., Datry,
T., Langhans, S.D. 2022. The terrestrial and semi-aquatic invertebrates
of intermittent rivers and ephemeral streams. Biological Reviews 97:
1408–1425.
Stubbington, R., Paillex, A., England, J., Barthès, A.,
Bouchez, A., Rimet, F., Sánchez-Montoya, M.M., et
al. 2019. A comparison of biotic groups as dry-phase indicators of ecological
quality in intermittent rivers and ephemeral streams. Ecological Indicators 97: 165-174.
Suárez, M.L.,
Vidal-Abarca, M.R. 2008. Índice para
valorar el estado de conservación de las ramblas mediterráneas (Índice de
Alteración de Ramblas = IAR). Tecnología del Agua 293: 67-78.
Tapolczai, K., Selmeczy, G. B., Szabó, B., B-Béres, V., Keck,
F., Bouchez, A., Rimet, F., et al. 2021. The potential of exact sequence variants (ESVs) to
interpret and assess the impact of agricultural pressure on stream diatom
assemblages revealed by DNA metabarcoding. Ecological Indicators 122:
107-322.
Tonkin, J.D., Bogan, M.T. Bonada, N., Rios-Touma,
B., Lytle, D.A. 2017. Seasonality and predictability shape temporal species
diversity. Ecology 98(5): 1201-1216.
Tueros, I., Borja, A., Larreta, J., Rodríguez, J.G.,
Valencia, V., Millán, E. 2009. Integrating
long-term water and sediment pollution data in assessing chemical status within
the European Water Framework Directive. Marine Pollution Bulletin 58:
1389-1400.
Turrini, T., Dörler, D., Richter, A., Heigl, F.,
Bonn, A. 2018. The threefold potential of environmental citizen
science-Generating knowledge, creating learning opportunities and enabling
civic participation. Biological Conservation 225: 176-186.
Van den
Broeck, M., Waterkeyn, A.,
Rhazi, L., Grillas, P., Brendonck, L. 2015. Assessing the ecological integrity
of endorheic wetlands, with focus on Mediterranean temporary ponds. Ecological
Indicators 54: 1-11.
Vander
Vorste, R., Stubbington,
R., Acuña, V., Bogan, M.T., Bonada, N., Cid, N., Datry, T., et al. 2021.
Climatic aridity increases temporal nestedness of invertebrate communities in
naturally drying rivers. Ecography 44: 860-869.
Vidal-Abarca, M.R., Gómez, R., Sánchez-Montoya,
M.M., Arce, M.I., Nicolás, N., Suárez, M.L. 2020. Defining Dry Rivers as the Most
Extreme Type of Non-Perennial Fluvial Ecosystems. Sustainability 12: 7202.
Vidal-Abarca, M.R., Nicolas-Ruiz, N., Sánchez-Montoya, M.M.,
Suárez, M.L. 2023. Ecosystem
services provided by dry river socio-ecological systems and their drivers of
change. Hydrobiologia 850: 2585–2607.
Vieira, C., Aguiar, F.C., Portela, A.P., Monteiro, J., Raven,
P.J., Holmes, N.T.H., Cambra, J., et al. 2018. Bryophyte communities of Mediterranean Europe: a first
approach to model their potential distribution in highly seasonal rivers. Hydrobiologia
812: 27–43.
Vinebrooke, R.D., Cottingham, K.L., Norberg, M.S.J,
Dodson, S.I, Maberly, S.C., Sommer, U. 2004. Impacts of multiple stressors on
biodiversity and ecosystem functioning: the role of species co-tolerance. Oikos
104: 451-457.
Westwood, C.G., England, J., Hayes,
C., Johns, T., Stubbington, R. 2021. The Plant Flow Index: A new method to
assess the hydroecological condition of temporary rivers and streams. Ecological
Indicators 120: 106964.
When, U., Rusca, M., Evers, J., Lanfranchi, V.
2015. Participation in flood risk management and the potential of citizen
observatories: a governance analysis. Environmental
Science and Policy 48: 225-236.