Introducción
Las arañas del género Latrodectus (Walckenaer, 1805)
son de gran interés sanitario debido a la presencia de una neurotoxina llamada
α-Latrotoxina, que puede causar la muerte en casos extremos, tanto en humanos
como en animales domésticos (de Roodt et al. 2017).
En la actualidad, el género Latrodectus está compuesto por 34 especies (World Spider Catalog 2023) que presentan
una amplia distribución en diferentes continentes e islas oceánicas. Ábalos (1980) cita para Argentina siete especies de
este género, L. geometricus (Koch,
1841), L. antheratus (Badcock, 1932), L. variegatus (Nicolet,
1849), L. diaguita (Carcavallo, 1960), L. corallinus (Abalos,
1980), L. quartus (Abalos, 1980) y L. mirabilis (Holmberg, 1876).
De estas especies, L. mirabilis es la de mayor distribución y la más
austral, ocupando diversas provincias argentinas, así como Uruguay (Capocasale y Pereira 2003) y el sur de Brasil
(Ott et al. 2014). González
(1976, 1977, 1979)
y Schnack y González (1978) describieron
el ciclo de vida de las poblaciones de L. mirabilis en Sierra de la
Ventana, Argentina, detallando un ciclo reproductivo anual con adultos
presentes durante los meses cálidos (diciembre a abril). Los machos adultos
mueren poco después de la cópula, mientras que las hembras adultas mueren a
partir de marzo-abril. Las ootecas permanecen cerradas durante todo el
invierno, y en primavera comienza la dispersión de los juveniles, quienes pasan
por tres o cuatro mudas hasta alcanzar la edad adulta. Las ootecas de L.
mirabilis constan de tres capas de telas, con una punta característica en
uno de sus extremos (Ábalos 1962, 1980; González 1976), lo
que las distingue de otras ootecas de esta zona serrana (Giambelluca et al. 2019).
Sierra de la Ventana, según Arana et
al. (2021) pertenece a la Región Neotropical, Provincia Pampeana, Distrito
Pampeano Austral, con altitudes que varían desde los 250 hasta los 1239 m s. n.
m. Su flora y fauna están estrechamente relacionadas con las regiones del sur
(Subregión Patagónica), del oeste (Provincia del Monte) y principalmente con la
provincia que la contiene que es la Provincia Pampeana (Guerrero y Apodaca 2022). En esta sierra,
predominan los pastizales naturales serranos (Bilenca
y Miñarro 2004) y se destacan por albergar una gran diversidad de especies
y un alto grado de endemismos (Guerrero y
Apodaca 2022). En esta zona, Frangi y
Bottino (1995) llevaron a cabo una evaluación integral de las comunidades
vegetales en función de la orientación de la ladera, la pendiente, la altitud y
el suelo, identificando 16 unidades vegetales (UV). Lizzi
et al. (2007) basándose en las UV del trabajo de Frangi y Bottino (1995) realizaron un mapeo de
las comunidades vegetales utilizando datos de campo combinando imágenes
multiespectrales, con variables geo-edafológicas, altitud, orientación y
pendiente.
Comprender la distribución de las especies en el espacio y
el tiempo, es un aspecto fundamental en el campo de la ecología. Esto ha
impulsado el desarrollo de los modelos de distribución de especies (SDM, por
sus siglas en inglés), donde se basan en la premisa de que los individuos de
una especie se distribuyen de acuerdo con patrones específicos que pueden ser
relacionados con variables ambientales predictivas, a fin de evaluar los
patrones de presencia de especies y la idoneidad de su hábitat. Los SDM se han
convertido en un componente clave en diversos campos de estudio, como la
ecología, la biología evolutiva, el manejo de la vida silvestre y la
conservación. Su amplio uso se debe a su eficacia y versatilidad para analizar
datos de ocurrencia y predecir la idoneidad del hábitat. Los factores
ambientales tienen un efecto sobre la distribución de las especies, ya sea de
una forma directa o indirecta (Guisan y
Zimmermann 2000). Estas relaciones entre organismos y medio abiótico son
una de las causas de los patrones espaciales de distribución (Mateo et al. 2011). Para los SDM se utilizan dos tipos
de información. Por un lado, los datos de las observaciones directas de la
especie de interés, que pueden obtenerse a través de técnicas de recolección en
el campo o mediante el acceso a bases de datos de colecciones científicas, los
que proporcionan información fundamental sobre la presencia y localización de
la especie en estudio. Por otro lado, las variables predictivas, que se
refieren a información relacionada con el clima, topografía o el entorno
ambiental de la zona de interés, las cuales se obtienen a través de sensores
remotos. A partir de programas estadísticos, se combinan estos datos para
predecir una distribución potencial de una especie en función de las
características del entorno. Existen trabajos sobre la aplicación de SDM de Latrodectus
sp. a grandes escalas regionales, utilizando base de datos de presencias con
imágenes de baja resolución y variables climáticas derivadas de modelos como el
WorldClim (Taucare-Ríos et al. 2016). Sin
embargo, no hay estudios a escalas locales utilizando datos de muestreos y de
imágenes de resolución media-alta que establecerían una determinación de su distribución
más precisa. Los ambientes protegidos, como reservas o parques de conservación
de la biodiversidad, son lugares ideales para realizar este tipo de estudio, ya
que la influencia antrópica se encuentra minimizada.
El presente estudio tiene como objetivo identificar sitios
con presencia de individuos adultos y ootecas de L. mirabilis en el
Parque Provincial E. Tornquist (Sierra de la Ventana, Buenos Aires), analizar su
distribución a través de las UV descritas por Lizzi et
al. 2007 y generar un modelo de distribución de especie (SDM) para L.
mirabilis, mediante la utilización de sensores remotos y datos recolectados
en el Parque. Se espera que estos resultados contribuyan al conocimiento de L.
mirabilis y su preferencia de hábitat; proporcionando información relevante
para la conservación de estos ecosistemas serranos y tener mapas predictivos de
encuentro de esta especie. Conocer la distribución de L. mirabilis a
escala local, permitirá, además, abordar aspectos relacionados con su ecología
y generar mapas de interés sanitario.
Materiales y métodos
Muestreo
El relevamiento de los sitios (SIT) con presencia de
individuos adultos y ootecas de L. mirabilis se llevó a cabo en el
Parque Provincial E. Tornquist, ubicado en Sierra de la Ventana (Buenos Aires,
Argentina), que tiene una extensión de 6718 hectáreas. La metodología utilizada
fue observación directa sin colecta, a partir de transectas (track) al azar. Se
realizaron siete campañas recorriendo un total de 56.87 km comprendidos en las
distintas transectas (Fig. 1). Las observaciones
se llevaron a cabo desde diciembre de 2022 hasta febrero de 2023, entre las
7:30 a.m. y las 3:30 p.m. Se georreferenciaron los sitios y se registró la
altitud de cada uno utilizando un dispositivo GPS Garmin, se tomaron
fotografías digitales del lugar y se registró la presencia o ausencia de rocas
en el sitio hallado. En cada transecta, se determinó una distancia de
observación de 5 m hacia cada lado. Para obtener la superficie recorrida se
multiplicó la longitud de la transecta por el ancho (10 m) y se expresó en
hectáreas (ha).
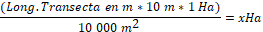
Análisis de densidad de hallazgos
Para el análisis de densidad de los registros fue utilizado
el programa QGIS, en el cual se trabajó con las unidades vegetales (UV) del
trabajo de Lizzi et al. (2007) en el cual, a los
archivos vectorial de líneas de los track, se les aplicó un buffer de distancia
fija de un segmento de 5 m, obteniendo un archivo de polígono, el cual se
combinó con la capa de UV mediante la herramienta de intersección de polígonos.
Con la calculadora de campos se cuantificó el área de cada UV y la inversa de
la misma se multiplicó por la cantidad de hallazgos obteniendo la densidad de SIT
por ha.
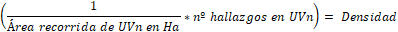
También se calculó la disponibilidad de UV en el Parque
Provincial E. Tornquist y se la cotejó con el área recorrida mediante una
comparación de medias (test t student con un P=0.05).
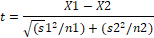
Modelo de distribución de especies
El modelo de distribución de especies (SDM) para L.
mirabilis comprendió dos pasos, utilizando la plataforma Google Earth
Engine (GEE). GEE es una plataforma que ofrece análisis geoespaciales de
alto rendimiento al aprovechar el procesamiento en la nube de Google y una
amplia colección de conjuntos de datos. Su catálogo cuenta con una variedad
diversa de datos geoespaciales, como imágenes satelitales de alta resolución,
así como información climática, topográfica y de vegetación. Esto permite
manejar grandes volúmenes de datos de manera ágil y precisa (Gorelick et al. 2017; Crego
et al. 2022).
El primer paso del modelo de distribución de especies fue
seleccionar las variables predictivas que permiten comprender y predecir la distribución
de L. mirabilis. Este proceso implicó la generación de múltiples
variables relacionadas como diversos índices de vegetación, características
geomorfológicas del entorno y datos de temperatura. Una vez generadas las
variables, se procedió a extraer los valores correspondientes a cada una de
ellas en los sitios de muestreo relevantes. Posteriormente, se utilizó el software
R (https://www.R-project.org/) para
construir una matriz de correlaciones, donde se seleccionaron las variables más
relevantes, que no estuvieran fuertemente correlacionadas entre sí (Araújo et al. 2019).
El segundo paso, fue generar el modelo de distribución
potencial. Para dicho modelo se utilizó la técnica de Random Forest
(RF), que consiste en un conjunto de árboles de clasificación y regresión
(CART) basados en el método de agregación bootstrap (bagging). Se trata
de un método no paramétrico y robusto (Hastie et al.
2001). En este estudio se empleó el 70% de los datos censados para
construir el modelo, junto con las variables ambientales seleccionadas y el 30%
de datos censados restante, fueron utilizados para evaluar el modelo. El
algoritmo se ejecuta 100 veces, generando así 100 modelos diferentes.
Posteriormente se valida la sensibilidad del modelo, para ello se obtienen los
valores de sensibilidad y especificidad de la curva ROC (Receiver Operating
Curve). La curva ROC es una representación gráfica de la capacidad discriminativa
de un modelo en diferentes puntos de corte. Se utiliza con datos de
presencia/ausencia y muestra el error de omisión en el eje de ordenadas
(clasificar una presencia como ausencia) y el error de comisión en el eje de
abscisas (clasificar una ausencia como presencia). El área bajo la curva ROC
(AUC) es equivalente al estadístico U de Mann-Whitney y representa la
probabilidad de que el modelo clasifique correctamente un par de casos de
presencia y ausencia. Un valor de AUC entre 0 y 1 indica la calidad del modelo,
donde 1 representa una clasificación perfecta, 0.5 indica clasificación
aleatoria y valores menores a 0.5 indican una clasificación peor que el azar (Mateo et al. 2011, Sofaer et
al. 2019). Para la presentación de los mapas se utilizó QGIS (versión
3.16).
Resultados
En las siete campañas se lograron 61 registros de sitios (Fig. 1): 24 con una ooteca cada uno, siete con una ooteca
y una hembra adulta cada uno, uno con una ooteca y un macho adulto, uno con dos
hembras adultas juntas, 24 con una hembra adulta, uno con un macho y una hembra
adultas, dos con un macho adulto cada uno y uno con una hembra subadulta.
Los hallazgos de los SIT se hicieron a distintas altitudes,
siendo el rango de 350 a 550 m s. n. m. el de mayor presencia (45), seguido por
el de 551 a 750 m s. n. m. (13), por el de 751 a 950 m s. n. m. (1) y por el de
951 a 1150 m s. n. m. (2). En todos los SIT se observaron afloramientos
rocosos.
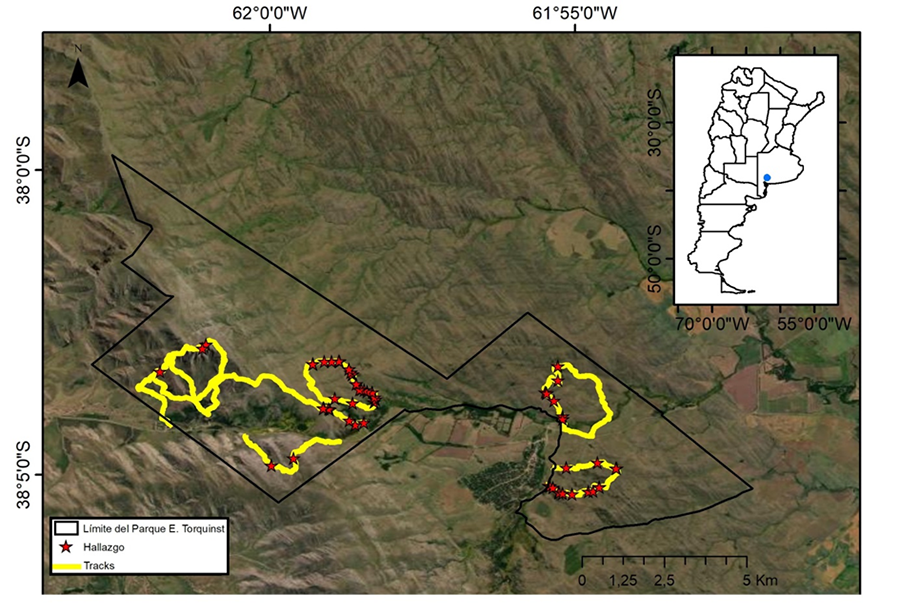
Figura 1. Mapa de transectas (Tracks) y
hallazgos de sitios con individuos adultos y ootecas de L. mirabilis en
el Parque Provincial E. Tornquist (Buenos Aires, Argentina).
Figure 1. Map of transects (Tracks) and findings of siteswith adult individual and egg sac of L.
mirabilis in the E. Tornquist Provincial Park
(Buenos Aires, Argentina).
La superficie recorrida de las transectas fue de 56.87 ha las
que pasaron por todas las UV a excepción de la UV 5, lo cual no es significativo
porque sólo comprende 2.86 ha de todo el Parque Provincial E. Tornquist. Todas
las demás UV fueron bien representadas y mediante el test de Student (P=0.05),
se comprobó que no hubo diferencias significativas entre la superficie de las
16 UV de Lizzi et al. (2007) del Parque Provincial
E. Tornquist y las exploradas en este estudio (Tabla 1).
Las UV que tuvieron mayor presencia de SIT fueron la 6, 7, 8 y 9, el resto de
las UV mostro menos de seis SIT (Tabla 1). En
cuanto a la densidad de SIT se observó una media de 1.07 sitios por
hectárea (Fig. 2), siendo la UV 11 la de mayor
densidad, seguida por la 9, 6, 7, 12, 15, 14, 8, 13, 3 y 2 (Tabla
1). Al cotejar la disponibilidad de UV en el Parque Tornquist (Media
491.42; Desvío 397.95) con las áreas recorridas (Media 355; Desvío 3.43) se
obtuvo un P= 0.0002.
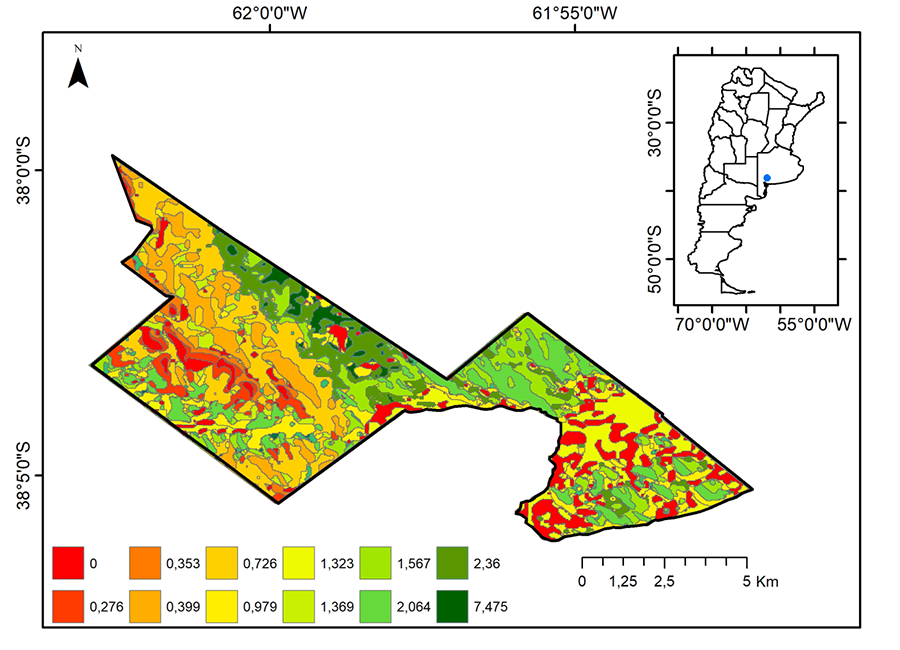
Figura 2. Densidad de sitios con individuos
adultos y ootecas de L. mirabilis en cada UV del Parque Provincial E.
Tornquist (Buenos Aires, Argentina).
Figure 2. Density of the sites with
adult individual and egg sac of L. mirabilis in
each UV of the E. Tornquist Provincial Park (Buenos Aires, Argentina).
Tabla 1. Hallazgos y
densidad de sitios con individuos adultos y ootecas de L. mirabilis en
las Unidades Vegetales (UV) descriptas por Lizzi et al.
2007, del Parque E. Tornquist (Buenos Aires, Argentina).
Table
1. Findings and density of the sites with adult individual and egg sac of L. mirabilis in the Vegetation Units
(VU) described by Lizzi et al.
2007, from E. Tornquist Park (Buenos Aires,
Argentina).
Modelo de distribución
De las nueve variables ambientales calculadas a partir de
distintos sensores y modelos disponibles en el catálogo de GEE, se
seleccionaron las siguientes cinco variables, que muestran alta correlación
entre los valores de presencia-ausencia y baja correlación entre ellas:
·
Valores medios de índice NDVI (Normalized Difference Vegetation
Index) y GLI (Green Leaf Index), de los meses de verano del 2022-2023. Para
ambos índices se utilizaron imágenes de Sentinel 2 de resolución 10 metros,
libre de nubes. El NDVI indica el vigor fotosintético de la vegetación. Se
calcula a partir de la diferencia normalizada entre la reflectancia en el
infrarrojo cercano (NIR) y la reflectancia en el rojo (RED) de una superficie
terrestre. El Índice GLI se calcula utilizando las bandas de rojo (RED), verde
(GREEN) y azul (BLUE) de la imagen y permite discriminar zonas vegetales (Vsrnr et al. 2022).
·
Mapas de pendiente y orientación, a partir de un modelo de
elevación de 30 metros provisto por la NASA. Se calcula la pendiente y la
orientación, a partir de un Modelo Digital de Elevación (DEM) de 30 m de
resolución. La pendiente o inclinación de una superficie en un determinado
punto, se calcula como el cambio vertical de elevación dividido por la
distancia horizontal. La orientación o dirección de la pendiente en un punto
determinado del DEM, se expresa en grados y muestra hacia qué dirección se inclina
una superficie. Ambas son medidas importantes para el análisis topográfico y la
caracterización de un terreno.
·
Temperatura media de superficie o LST (Land Surface Temperature)
de los meses de verano del 2022-2023, a partir de Landsat 9, con resolución 30
m (Meng et al. 2022).
Teniendo en cuenta estas cinco variables surge el modelo de
distribución de especies propuesto para L. mirabilis en el Parque
Provincial E. Tornquist. Este modelo fue sensible a la heterogeneidad de
ambientes presentes, mostrando un valor de ajuste de 0.74 (AOC-ROC) (Fig. 3).
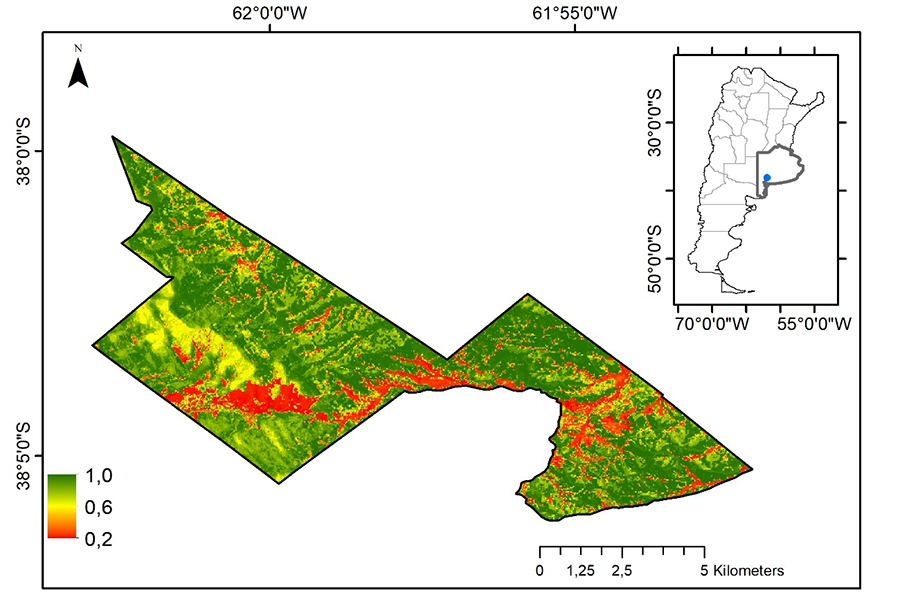
Figura 3. Modelo de distribución de especies
para L. mirabilis con valores expresados en probabilidades para el
Parque Provincial E. Tornquist (Buenos Aires, Argentina).
Figure 3. Species distribution model for L. mirabilis with values
expressed in probabilities for E. Tornquist Provincial Park (Buenos Aires,
Argentina).
Conclusión
La gran heterogeneidad vegetal y de relieve del Parque
Provincial E. Tornquist permite encontrar distintas abundancias de sitios
propicios para L. mirabilis según las combinaciones de las diferentes
condiciones microambientales, lo cual demuestra que la especie en estudio tiene
preferencias de hábitat. En cuanto a la altitud los hallazgos más frecuentes de
L. mirabilis estuvieron registrados desde los 368 a los 1125 m s. n. m.,
lo que indica que la altitud no fue una limitante para su presencia, si bien la
mayoría de los registros se obtuvieron entre los 350 y los 550 m s. n. m.
Tanto por el análisis con las UV o con los SDM ambos
tuvieron las mismas zonas como lugares posibles de encuentro, pudiéndose
concluir que L. mirabilis es una especie que prefiere pastizales bajos
que no superen los 10 cm, con afloramientos rocosos. El haber hallado una
densidad aproximada de 1.07 sitios/ha demuestra que el Parque Provincial E.
Tornquist es un lugar adecuado para esta especie y que algunas UV son mejores
que otras para su establecimiento.
El modelo de distribución de especie (SDM) para L.
mirabilis ajustó adecuadamente para la fecha del estudio y la metodología
usada de imágenes satelitales y su combinación da la posibilidad de hacer uso
de estos modelos en fechas prefijadas.
Si bien se ha realizado otro trabajo con temáticas parecidas
y hasta con este mismo género, pero con diferente especie, no son comparable
los resultados con los nuestros debido a que se ha utilizado el programa
Maxent, el cual trabaja en escala mayores a 30 km de lado, mientras que
nuestros resultados pueden ser usados a una escala mucho menor acorde a las
dimensiones del Parque estudiado. Pese a la gran importancia sanitaria de L.
mirabilis no se han realizado mapas de probabilidad de encuentro para esta
especie ni de densidad en zonas donde su presencia es conocida. Este trabajo
podría ser usado para la elección de zonas de caminatas donde se minimizaría el
riesgo de encuentro, si se toma en cuenta sus preferencias de hábitat y ciclo
biológico.
Contribución de los autores
LAG: aportó la idea original; conceptualización; colecta de
datos de campo; metodología; redacción (borrador original); supervisión; edición.
MdRI: investigación, visualización; redacción; metodología; software. AG:
investigación; validación; redacción; revisión.
Agradecimientos
A Áreas Protegidas de la Provincia de Buenos Aires por
permitirnos trabajar en el Parque, a todo el personal del mismo y a los
Guardaparques Diego Repetto y Cristian Peterson por facilitar el muestreo. Al
Dr. José M Lizzi por compartirnos los archivos shape de su trabajo.
Referencias
Ábalos, J.W. 1962. The Egg-Sac in the Identification of Species of Latrodectus
(Black-Widow Spiders). Psyche: A Journal of Entomology
69:268–270.
Ábalos, J.W. 1980. Las Arañas
del género Latrodectus en la Argenitna. Obra del Centenario del Museo
de La Plata 6:29–51.
Arana, M.D., Natale, E.,
Ferretti, N., Romano, G., Oggero, A., Posadas, P., Morrone, J.J., et al. 2021. Esquema
Biogeográfico de la República Argentina. Col. Opera Lilloana 56 (2021).
Fundación Miguel Lillo. Tucumán, Argentina. ISBN 978-950-668-039-8
Araújo, M.B., Anderson, R.P.,
Barbosa, A.M., Beale, C.M., Dormann, C.F., Early, R., Guisan, A., et al. 2019. Standards for distribution models in biodiversity assessments. Science
Advances 5(1):1–12. https://doi.org/10.1126/sciadv.aat4858
Bilenca, D., Miñarro, F.
2004. Identificacion de Areas Valiosas de Pastizal en las pampas y campos de
Argentina, Uruguay y sur de Brasil (AVPs). 1ra ed. Fundación Vida Silvestre
Argentina, Buenos Aires, Argentina. ISBN 950-9427-11-X.
Capocasale, R. M., Pereira,
A. 2003. Diversidad de la biota uruguaya. Araneae. Anales del Museo Nacional
de Historia Natural y Antropología 10:1–12. ISSN - 0797 -
6828
Crego, R.D., Stabach, J.A., Connette, G. 2022. Implementation of species
distribution models in Google Earth Engine. Diversity and
Distributions 28:904–916. https://onlinelibrary.wiley.com/doi/10.1111/ddi.13491
de Roodt, A.R., Lanari, L.C.,
Laskowicz, R.D., Costa de Oliveira, V., Irazu, L.E., González, A., Giambelluca,
L.A., et al. 2017. Toxicity of the venom of Latrodectus
(Araneae: Theridiidae) spiders from different regions of Argentina and
neutralization by therapeutic antivenoms. Toxicon 130:63–72. https://doi.org/10.1016/j.toxicon.2017.02.029
Frangi, J.L., Bottino,
O.J. 1995. Comunidades vegetales de la Sierra de la Ventana, Provincia de
Buenos Aires, Argentina. Revista de la Facultad de Agronomía 71:93–133.
Giambelluca, L.A.,
Giambelluca, A.L., González, A. 2019. Fauna acompañante intraooteca de Latrodectus
mirabilis (Araneae: Theridiidae) en Sierra de la Ventana, Buenos Aires,
Argentina. Historia Natural 9(1):53-62.
González, A. 1976.
Observaciones bioecológicas sobre una especie del género Latrodectus
(Walcknaer, 1805), del grupo mactans de Sierra de la Ventana (Provincia de
Buenos Aires, Argentina) (Araneae, Theridiidae). II Produccuon de huevos. Neotropica.
22:129–131.
González, A. 1977.
Observaciones bioecológicas sobre una especie del género Latrodectus
(Walcknaer, 1805), del grupo mactans de Sierra de la Ventana (Provincia de
Buenos Aires, Argentina) (Araneae, Theridiidae). I Hábitat y ciclo Vital. PHYSIS,
Sección C. 36:277–282.
González, A. 1979.
Observaciones bioecológicas sobre una especie del género Latrodectus
Walc- kenaer, 1805, del grupo mactans de Sierra de la Ventana (Prov. de Buenos
Aires, Argentina) (Araneae, Theridiidae). III Desarrollo
postembrionario. Acta Zoológica Lilloana 35:95-110.
Gorelick, N., Hancher, M., Dixon, M., Ilyushchenko, S., Thau, D., Moore,
R. 2017. Google Earth Engine: Planetary- scale
geospatial analysis for everyone. Remote Sensing of Environment
202:18–27. https://doi.org/10.1016/j.rse.2017.06.031
Guerrero,
E.L., Apodaca, M.J. 2022. The smallest area shaped a big problem: a revision of the placement
of the Ventania sky island in the biogeography of South America. Biological
Journal of the Linnean Society 20:1-16. https://doi.org/10.1093/biolinnean/blac088
Guisan,
A., Zimmermann, N.E. 2000. Predictive habitat
distribution models in ecology. Ecological Modelling 135:147-186. https://doi.org/10.1016/S0304-3800(00)00354-9
Hastie, T., Tibshirani, R., Friedman, J.H.
2001. The
elements of statistical learning: data mining, inference, and prediction: with
200 full-color illustrations. pp. 745.
Springer. ISBN 0-387-95284-5. OCLC 46809224.
Lizzi, J.M., Garbulsky, M.F., Golluscio, R.A., Deregibus, A.V. 2007. Mapeo indirecto de la vegetación de Sierra de la Ventana,
provincia de Buenos Aires. Ecologia Austral 17:217–230. versión On-line
ISSN 1667-782X
Mateo, R.G., Felicísimo, A.M.,
Muñoz, J. 2011. Modelos de distribución de especies: Una revisión sintética
Species distributions models: A synthetic revision. Revista Chilena de
Historia Natural 84:217-240. http://dx.doi.org/10.4067/S0716-078X2011000200008
Meng, X., Cheng, J., Guo, H., Guo, Y., Yao B. 2022. Accuracy Evaluation
of the Landsat 9 Land Surface Temperature Product. IEEE Journal of Selected
Topics in Applied Earth Observations and Remote Sensing 15:8694-8703. http://dx.doi.org/10.1109/JSTARS.2022.3212736
Ott, R., Nei, E., Rodrigues, L.
2014. First record of Latrodectus mirabilis (Araneae :
Theridiidae) from southern Brazil and data on natural history of the species. Revista
Colombiana de Entomología 40:305–310. ISSN 0120-0488
Schnack, J.A., González,
A. 1978. Observaciones bioecológicas sobre una especie del género Latro- dectus
Walckenaer, 1805, del grupo mactans de Sierra de la Ventana (Prov. de Buenos
Aires, Ar- gentina) (Araneae, Theridiidae). IV Producción anual. Neotrópica 24(71),
9-12.
Sofaer, H.R., Hoeting, J.A.,
Jarnevich, C.S. 2019. The area under the precision- recall
curve as a performance metric for rare binary events. Methods in Ecology and
Evolution 10(4):565–577. https://doi.org/10.1111/2041-
210X.1314
Taucare-Ríos, A., Bizama, G., Bustamante, R.O. 2016. Using
Global and Regional Species Distribution Models (SDM) to Infer the Invasive
Stage of Latrodectus geometricus (Araneae: Theridiidae) in the Americas. Environmental
Entomology 46(6): 1379-1385.
Vsrnr, R.T., Dua, S., Saha, P. 2022. Identifying Black Rice
Cultivated Area Using Sentinel 2. Journal of Scientific Research
66(2):214-219. https://doi.org/10.37398/JSR.2022.660228
World Spider Catalog 2023. World Spider Catalog. Version
24. Natural History Museum Berna, Suiza. [Consultado el 6 de junio de 2023]. http://wsc.nmbe.ch,
doi: 10.24436/2