Figure
3. Invasive and native threatened fish species from
the Cape Fold Ecoregion IRES. a) Example of invasive fish in South Africa: juvenile Micropterus
sp. (15 cm size); b)
Native South African species: Pseudobarbus afer (Peters, 1854) (IUCN Red
list status: endangered). Photo credits: D.J. Woodford.
Figura 3. Especies de peces invasoras y nativas
amenazadas de la ecorregión de Cape Fold IRES. a) Ejemplo de pez invasor
en Sudáfrica: juvenil Micropterus sp. (tamaño 15 cm); b) Especie
nativa sudafricana: Pseudobarbus afer (Peters, 1854) (Estado en la Lista
Roja de la UICN: en peligro de extinción). Créditos de las
fotos: D.J. Woodford.
Similarly,
intermittent reaches can act as refugia for native
invertebrate species and in some specific cases they may represent the only
portions of the river network which are free from IAS (e.g., intermittent
stretches only parts not invaded by New Zealand mudsnail Potamopyrgus
antipodarum in Californian creeks, Abramson 2009). In both perennial and
intermittent rivers, the most common invertebrate IAS are crustaceans and
molluscs, whereas aquatic insects are extremely rare IAS in both systems (Fenoglio et al. 2016; Burgazzi et
al. 2018). The multi-dimensionality and dynamism of
IRES habitat form an extra factor where fine scale spatial attributes are
important as they provide fractal micro-refugia which support even species with
no desiccation resistance traits (Burgazzi et al. 2018; Pardil et al. 2019). For instance, Procambarus clarkii tends to use complex
microhabitats (e.g., vegetated sections or under boulders) in ephemeral water
bodies of Portugal (Aquiloni
et al. 2005) and Kouba et al. (2016) emphasised the
evasive capacity of invasive crayfish (e.g., Procambarus virginalis and P.
clarkii) to dig vertical burrows into the hyporheic zone under drought
conditions.
Further, the drying regime and habitat
mosaic increase entrance of terrestrial invertebrate species into the community
in a manner that can transport propagules both
downstream, and laterally, with flow resumption (e.g., Rosado et al. 2015; Steward et al.
2017; Sánchez-Montoya et al. 2020; Sánchez-Montoya et al. 2023). Therefore, the incursion and timing of terrestrial and
semi-aquatic IAS is a topic which requires further research.
Encroachment of terrestrial IAS into IRES
is mostly characterised by vegetation which spreads along the riparian zone and
expands into newly available habitats formed in the emerging dry channels and
exposed riverbeds. During prolonged drought periods this may involve the entire
riverbed, with colonisation from the lateral riparian zone to the main riverbed
by generalist, pioneer and fast spreading vegetation on the exposed sediments
(e.g., Bolpagni
and DallaVecchia 2022). Drought resistant and
flood-adapted flora are then particularly favoured and the hydrologic dynamism
of IRES may assist spread through hydrochory. For instance, the Giant reed Arundo
donax, common IAS in numerous European and North American regions,
reproduce via asexual fragments (vegetative reproduction), and its spread is
accelerated through flooding events which transport fragments downstream
whereupon establishment occurs (Calazans et al. 2023; Ollero et al. 2022; Saltostall
et al. 2017, see Figs. 1 and 2).
Riparian
ecosystems of ephemeral streams in some areas of southern Africa (e.g.,
Namibia) have been affected by severe infestations of Prosopis spp.
that tend to form dense thickets. These are in some cases perceived as useful
for fodder and shade by farmers, but considered a critical threat to the
riparian forests displacing native vegetation and thus affecting the animal
taxa which utilise these habitats (Joubert 2009). Plant invasions (e.g.,
early stage) may tend to go unnoticed in arid areas (Milton and Dean 2010) also because of the close resemblance between arid-adapted alien
and some indigenous species (i.e., cryptic invasion processes: Morais and Reichard 2018). For instance, Australian Atriplex nummularia and Asian Tamarix
ramosissima are well adapted for dispersal in flooding rivers to mud banks
and floodplains, as are their indigenous congeners in Africa (Milton and Dean 2010). Moreover, alien riparian species like Tamarix spp., in the
USA and South Africa, are highly salt and desiccation tolerant which promotes
their capacity to colonise and stabilise floodplain soils, while their traits
ultimately affect channel morphology and hydrology (e.g., USA: Birken and Cooper 2006; Africa: Grenfell
and Dube 2022). Establishment of these species can
shift community traits from deciduous to evergreen species and may exacerbate
the drying phase by increasing transpiration rates (Zedler and Kercher 2004).
Knowledge to date, gaps and new
frontiers
A literature review using the Web of
Science publication database (April 25, 2023) combining terms related to
biological invasions and intermittent streams [(invas* OR alien OR non-native
OR exotic) AND (intermittent river OR ephemeral stream OR non-perennial water
way)] on the topic section (“title”, “abstract” and “keywords”) provided 112
results in English (110) and non-English (2) languages in a period from 1992 to
2023 (details in Tables S1-S2). Of these publications
57 somehow focussed on IAS and IRES and were mostly published after the year
2000. Approximately half of the publications concerned the fish community
(47.4%: 27 out of 57), followed by studies on vegetation (15) and invertebrates
(8). North America (32) and Europe (10) were the regions more explored, with
few publications based on Asia, Africa or South America. These outputs stress
that Global South and some target organisms (i.e., invertebrates and
microorganisms) are still underrepresented in the current international
scientific literature. This geographic pattern appears common in river science,
as seen in a recent review of stream communities and ecosystem functioning (Colls et al. 2024).
In this context, there has been relatively
little direct focus on the interplay between IRES and IAS due to a lack of
interdisciplinary research and quite likely as a result of unpredictable /
rapid wetting-drying events which are a barrier to extensive field expeditions
and to realistic/representative mesocosm experiments on large temporal and
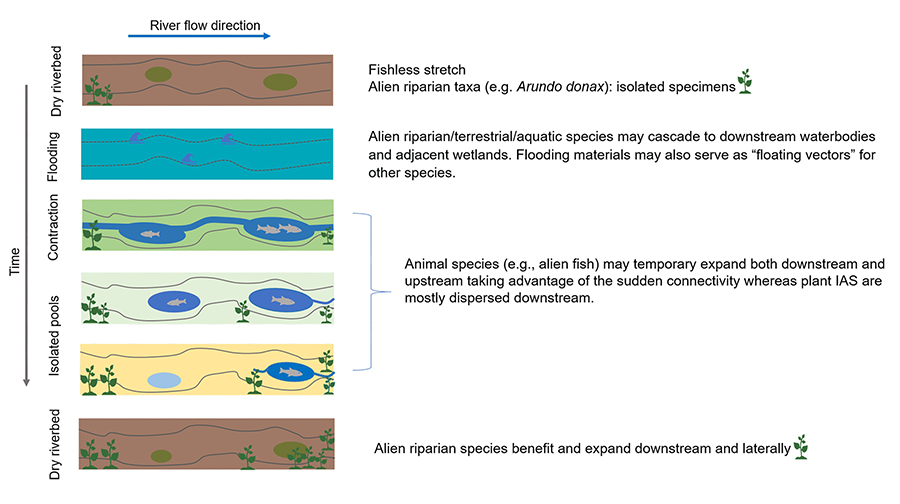
space scale. A general trend, however, is that during wet periods animal IAS
(e.g., fish) may migrate both downstream and upstream taking advantage of the
sudden connectivity whereas plant IAS (but theoretically also animals without
active dispersal) are mostly dispersed downstream in the flood period. Overall,
extreme persistent drought events prevent the arrival and spread of
water-dependent alien taxa. In fact, desiccation and water level management are
part of the aquatic IAS management toolbox, having been proposed to
eradicate/limit populations in artificial water bodies or channels (e.g., Paukstis et al. 1999).
Context
specific approaches specifically designed to consider communities and ecosystem
services in riverine systems are highly necessary to improve and enhance
research of biological invasions in flowing waters (Guareschi
et al. 2022; Mathers
et al. 2022), this
approach ought to be implicitly extended to IRES (e.g., biological invasion
hypotheses). There are various opportunities to exploit in developing the
interdisciplinary study of IAS and IRES as multiple stressors on biological
communities to understand and predict outcomes of global change, as well as
forming “natural arenas” for testing hypotheses in the field. For example, the
role of ecological matching and environmental filtering, where species with
broad physiological tolerances, fast generational turnover, life histories
linked to flood pulses or over-land linked dispersal traits, and generalist
consumers may be hypothesised to be more likely to invade IRES. In numerous
contexts most IAS (both plants and animals) already showed adaptations and life
histories which facilitate rapid, opportunistic exploitation of habitats and
resources (e.g., Zedler and Kercher 2004; Drown et al. 2011; Coverdale et al. 2013). Similarly, the nature of IRES as
transitional, ‘blank slate’, ecosystems may allow pioneering species to
colonise faster and restrict native species re-colonisation through predation
and niche exclusion. At the same time, the biotic resistance hypothesis (Kennedy
et al. 2002) can be
also tested in IRES where the native assemblage is pre-adapted to the local
hydrology to understand the mechanisms of invasion. In this context, it can be
hypothesized that the focal invasive species would exploit a gap in an
ecological niche specific to IRES rather than competitively excluding
pre-adapted native species from a given niche. Interdisciplinary ventures which
predict drying-wetting events can be used to target specific locations for
monitoring to jointly understand the drivers and processes behind colonisation
succession in invaded and uninvaded IRES, which can aid understanding of poorly
documented phenomena such as invasion meltdown (e.g. Braga et
al. 2020). A further
aside of interest is an assessment of the structure of interspecific
interactions and trophic pathways considering the shifts between terrestrial
dominated and aquatic dominated habitats in the same mosaic. These systems
should be also studied comparatively at a global scale to understand the
linkages in systems adapted to a flood pulse, such as Afro and Neo tropical
wetlands. However, there are considerable challenges in the successful
implementation of before-after-control-impact approaches due to their high
hydrological dynamism.
Physiological preference and desiccation
resistance should be further investigated, ideally at different population
levels, especially for successful IAS (e.g., Guareschi and Wood 2020) to increase our knowledge and understanding on future potential
invaders in IRES. At the same time emerging monitoring approaches like
eDNA-based species detections showed promising outputs from isolated pools in
IRES, especially for fish communities (Bylemans et al. 2016). Nonetheless,
technical development and standardised methodology needs to be completed to
fully understand and exploit eDNA detection in hydrologically variable systems.
For example, the degradation of eDNA and downstream transport of eDNA during a
flash drying or flooding event, and the capacity to detect historical
communities from dry reach sediment. Finally, the implications of biological
invasions and naturally intermittent hydrological regimes should be used to
predict outcomes of climate induced changes to flow regimes in historically
perennial systems now threatened by drought and abstraction (e.g., Alpine
streams: Piano et al. 2019). Further advances in trait based predictive ecology, combined with
international cohesion, has the potential to advance the knowledge regarding
management of IAS and IRES at a global scale.
Acknowledgements
Authors wish to thank the editors and
reviewers of this special issue. SG was supported by the project "SUMHAL -
Sustainability for Mediterranean Hotspots integrating LifeWatchERIC",
funded by: MICINN through European Regional Development Fund [SUMHAL,
LIFEWATCH-2019-09-CSIC-13, POPE 2014-2020] at Doñana Biological Station
(EBD-CSIC).
Contribution of the authors
Simone Guareschi, Josie South:
Conceptualisation, Methodology, Writing - Review and editing.
References
Abramson, M., 2009. Tracking the invasion
of the New Zealand mudsnail, Potamopyrgus antipodarum, in the Santa
Monica Mountains. Urban Coast 1, 21–27.
Aquiloni, L., Ilhéu, M., Gherardi, F. 2005. Habitat use and dispersal of the invasive
crayfish Procambarus clarkii in ephemeral water bodies of Portugal. Marine
and Freshwater Behaviour and Physiology 38(4), 225-236.
Arthington,
A.H., Balcombe, S.R. 2011. Extreme flow variability
and the ‘boom and bust’ecology of fish in arid‐zone
floodplain rivers: a case history with implications for environmental flows,
conservation and management. Ecohydrology 4(5), 708-720.
Belmar, O., Bruno, D., Guareschi,
S., Mellado‐Díaz, A.,
Millán, A., Velasco, J., 2019. Functional responses of aquatic
macroinvertebrates to flow regulation are shaped by natural flow intermittence
in Mediterranean streams. Freshwater Biology 64(5), 1064-1077.
Birken, A.S.,
Cooper, D.J. 2006. Processes of Tamarix
invasion and floodplain development along the lower Green River, Utah. Ecological
Applications 16(3), 1103-1120.
Bolpagni,
R., Dalla Vecchia, A. 2022. A longitudinal snapshot
of pioneer plant patterns along lowland temperate rivers. River Research and
Applications 38(6), 1129-1138.
Braga, R.R.,
Ribeiro, V.M., Padial, A.A., Thomaz, S.M., Affonso, I.D.P., Wojciechowski, J.,
et al. 2020. Invasional meltdown: an experimental
test and a framework to distinguish synergistic, additive, and antagonistic
effects. Hydrobiologia 847, 1603-1618.
Broom, C.J., Weyl,
O.L.F., South, J. 2023. Fish community, stressors
and conservation in the Rondegat River (South Africa). Fishes in
Mediterranean Environments 2023.001: 21p. https://doi.org/10.29094/FiSHMED.2023.001
Bruno, D., Hermoso, V., Sánchez‐Montoya, M.M., Belmar, O.,
Gutiérrez‐Cánovas, C.,
Cañedo‐Argüelles, M. 2022.
Ecological relevance of non‐perennial
rivers for the conservation of terrestrial and aquatic communities. Conservation
Biology, e13982.
Bruton, M.N. 1979. The survival of habitat desiccation by air breathing clariid
catfishes. Environmental Biology of Fishes 4, 273-280.
Burgazzi,
G., Guareschi, S., Laini, A., 2018. The role of
small-scale spatial location on macroinvertebrate community in an intermittent
stream. Limnetica 37(2), 319-340.
Bylemans, J.,
Furlan, E.M., Pearce, L., Daly, T., Gleeson, D.M. 2016. Improving the containment of a freshwater invader using
environmental DNA (eDNA) based monitoring. Biological Invasions 18,
3081-3089.
Calazans, E., Lopes, A., Girotto, L., de Paula, A.L.O., Franco, A.C.,
Ferreira, C.S. 2023. Mechanical control inadvertently
increases risk of alien plant invasion: influence of stem fragmentation and
inundation regimes on Arundo donax regeneration in Neotropical savanna. Australian
Journal of Botany 71(5) 223-230.
Chapman, D.S., Gunn, I.D., Pringle, H.E., Siriwardena, G.M., Taylor, P.,
Thackeray, S.J., Willby, N.J. and Carvalho, L., 2020. Invasion of freshwater
ecosystems is promoted by network connectivity to hotspots of human activity. Global
Ecology and Biogeography 29(4), 645-655.
Chiu, M.C., Leigh, C., Mazor, R., Cid, N., Resh, V. 2017. Anthropogenic
threats to intermittent rivers and ephemeral streams. In: Datry, T., Bonada, N., Boulton, A.J. (Eds.), Intermittent
Rivers and Ephemeral Streams, pp. 433–454. Academic Press. https://doi.org/10.1016/B978-0-12-803835-2.09997-6
Clavero, M., Esquivias, J.,
Qninba, A., Riesco, M., Calzada, J., Ribeiro, F., Fernández, N., Delibes, M.
2015. Fish invading deserts: non‐native
species in arid Moroccan rivers. Aquatic Conservation: Marine and Freshwater
Ecosystems 25(1), 49-60.
Colls, M., Viza, A., Zufiarre, A., Camacho-Santamans, A., Laini, A.,
González Ferreras, A.M., et al. 2024. Impacts of diffuse urban stressors on
stream benthic communities and ecosystem functioning: A review. Limnética
43(1), 89-108. https://doi.org/10.23818/limn.43.07
Cottet, M., Robert, A., Tronchère-Cottet, H., Datry, T. 2023. It's dry, it
has fewer charms!: Do perceptions and values of intermittent rivers interact
with their management?. Environmental Science & Policy 139, 139-148.
Corti, R.,
Datry, T. 2012. Invertebrates and sestonic matter
in an advancing wetted front travelling down a dry river bed (Albarine,
France). Freshwater Science 31(4), 1187-1201.
Coverdale, T.C., Axelman, E.E., Brisson, C.P., Young, E.W., Altieri, A.H.,
Bertness, M.D. 2013. New England salt marsh recovery: opportunistic
colonization of an invasive species and its non-consumptive effects. PLoS
One 8(8), p.e73823.
Datry, T., Larned, S.T., Tockner, K. 2014. Intermittent rivers: a
challenge for freshwater ecology. BioScience 64(3), 229-235.
Datry, T., Bonada, N., Boulton, A.J. 2017. Intermittent Rivers and Ephemeral Streams: Ecology
and Management. Academic Press, Elsevier, 597 pp.
Drown, D.M., Levri, E.P., Dybdahl, M.F. 2011. Invasive genotypes are
opportunistic specialists not general purpose genotypes. Evolutionary
Applications 4(1), 132-143.
Ellender, B.R., Weyl, O.L.F., Swartz, E.R. 2011. Invasion of a headwater
stream by non-native fishes in the Swartkops River system, South Africa. African
Zoology 46:39–46
Ellender, B.R., Woodford, D.J., Weyl, O.L.F. 2015. The invasibility of small
headwater streams by an emerging invader, Clarias gariepinus. Biological
Invasions 17, 57–61.
Ellender, B.R., Weyl, O.L.F., Alexander, M.E., Luger, A.M., Nagelkerke,
L.A.J., Woodford, D.J. 2018. Out of the pot and into the fire: Explaining the
vulnerability of an endangered small headwater stream fish to black‐bass
Micropterus spp. invasion. Journal of fish biology 92(4),
1035-1050.
Fenoglio, S., Bonada, N.,
Guareschi, S., López-Rodríguez, M.J., Millán, A., Tierno de Figueroa, J.M.
2016. Freshwater ecosystems and aquatic insects: a paradox in
biological invasions. Biology letters 12(4), p.20151075.
Firmat, C., Alibert, P., Losseau, M., Baroiller, J. F., Schliewen, U.K.
2013. Successive invasion-mediated interspecific hybridizations and population
structure in the endangered cichlid Oreochromis mossambicus. PLoS One,
8(5), e63880.
Francis,
R.A., Chadwick, M.A. 2012. Invasive alien species
in freshwater ecosystems: a brief overview. In: Francis, R.A. (Ed.) A
handbook of global freshwater invasive species, pp. 23-41. Earthscan, New
York, USA.
Grenfell,
M.C., Dube, T. I. 2022. Spectral classification,
mapping and physical habitat implications of a riparian invasion by Tamarix
ramosissima Ledeb. in the Touws River, Klein Karoo, South Africa. African
Journal of Aquatic Science, 47(2), 197-206.
Guareschi,
S., Wood, P.J. 2020. Exploring the desiccation
tolerance of the invasive bivalve Corbicula fluminea (Müller 1774) at
different temperatures. Biological Invasions 22, 2813–2824.
Guareschi,
S., Wood, P.J. 2022. Biological Invasions of River
Ecosystems: A Flow of Implications, Challenges, and Research Opportunities. In:
Dominick, A., DellaSala, M.I., Goldstein, I. (Eds.), Imperiled: The
Encyclopedia of Conservation, Elsevier, 485-498, https://doi.org/10.1016/B978-0-12-821139-7.00147-1
Guareschi,
S., Wood, P.J., England, J., Barrett, J., Laini, A. 2022. Back to the future: Exploring riverine macroinvertebrate
communities' invasibility. River Research and Applications, 38(8), 1374-1386.
IPBES 2023. Summary for Policymakers of the Thematic Assessment Report on
Invasive Alien Species and their Control of the Intergovernmental
Science-Policy Platform on Biodiversity and Ecosystem Services. Roy, H.E.,
Pauchard, A., Stoett, P., Renard Truong, T., Bacher, S., Galil, B.S., Hulme,
P.E., et al. (eds.). IPBES secretariat, Bonn, Germany. https://doi.org/10.5281/zenodo.7430692
Joubert, D.F. 2009. Invasive plants in Namibian subtropical and riparian woodlands.
In: Koli, R.K., Jose, S., Singh, H.P., Batish, D.R. (Eds.), Invasive Plants
and Forest Ecosystems. Chapter 20. CRC press, Francis and Taylor Group,
Boca Raton, FL, USA. https://doi.org/10.1201/9781420043389
Kennedy, T.A., Naeem, S., Howe, K.M., Knops, J.M., Tilman, D., Reich, P.
2002. Biodiversity as a barrier to ecological invasion. Nature 417,
636-638.
Kerezsy, A., Gido, K., Magalhães, M.F., Skelton, P.H. 2017. The biota of
intermittent rivers and ephemeral streams: Fishes. In: Datry, T., Bonada, N., Boulton, A.J. (Eds.). Intermittent rivers and ephemeral
streams, pp. 273-298. Academic Press.
Kouba, A., Tíkal, J., Císař, P., Veselý, L., Fořt, M., Příborský, J.,
Patoka, J., Buřič, M. 2016. The significance of droughts for hyporheic
dwellers: evidence from freshwater crayfish. Scientific Reports 6(1),
26569.
Mathers, K.L., Guareschi, S., Pattison, Z. 2022. Biological invasions in
rivers and associated ecosystems: New insights, challenges, and methodological
advancements. River Research and Applications 38(8), 1351-1355.
Messager, M.L., Lehner, B., Cockburn, C., Lamouroux, N., Pella, H., Snelder,
T., Tockner, K., et al. 2021. Global prevalence of non-perennial rivers and
streams. Nature 594, 391–397. https://doi.org/10.1038/s41586-021-03565-5
Milton, S.J.,
Dean, W.R.J. 2010. Plant invasions in arid areas:
Special problems and solutions: A South African perspective. Biological
Invasions 12, 3935–3948.
Morais, P.,
Reichard, M. 2018. Cryptic invasions: A review. Science
of the Total Environment 613, 1438-1448.
Moyle, P.B.,
Light, T. 1996. Fish invasions in California: do
abiotic factors determine success. Ecology 77(6), 1666-1670.
Ollero, A., Conesa, C., Vidal-Abarca, M.R. 2022. A guide to good practices for the management and restoration of
mediterranean ephemeral streams: Resilience and adaptation to climate change. EDITUM, Servicio de Publicaciones de la Universidad de
Murcia, Murcia, España. 184 pp. ISBN: 978-84-09-33842-9.
Pařil, P., Polášek, M.,
Loskotová, B., Straka, M., Crabot, J., Datry, T. 2019. An
unexpected source of invertebrate community recovery in intermittent streams
from a humid continental climate. Freshwater Biology 64, 1971– 1983.
Paukstis, G.L., Tucker, J.K., Bronikowski, A.M., Janzen, F.J .1999.
Survivorship of aerially-exposed zebra mussels (Dreissena polymorpha)
under laboratory conditions. Journal of Freshwater Ecology 14, 511–517
Petitjean,
M.O.G., Davies, B.R. 1988. Ecological impacts of
inter-basin water transfers: some case studies, research requirements and
assessment procedures in southern Africa. South African Journal of Science
84(10), 819-828
Piano, E., Doretto, A., Falasco, E., Fenoglio, S., Gruppuso, L., Nizzoli,
D., Viaroli, P., et al. 2019. If Alpine streams run dry: the drought memory of
benthic communities. Aquatic Sciences 81, 1-14.
Planty‐Tabacchi, A.M., Tabacchi, E., Naiman, R.J., Deferrari, C., Decamps, H. 1996.
Invasibility of species‐rich communities in riparian zones. Conservation Biology
10(2), 598-607.
Rodríguez-Lozano,
P., Woelfle-Erskine, C., Bogan, M.T., Carlson, S.M. 2020. Are
non-perennial rivers considered as valuable and worthy of conservation as
perennial rivers? Sustainability 12(14), 5782.
Rosado, J., Morais, M., Tockner, K. 2015. Mass dispersal of terrestrial
organisms during first flush events in a temporary stream. River Research
and Applications 31, 912–917.
Sánchez-Montoya, M.M., Guerrero-Brotons, M., Miñano, J., Gómez, R. 2020. Effects of
debris piles and pools along dry riverbeds on nutrients, microbial activity,
and ground-dwelling arthropods: a Namibian ephemeral river case. Journal of
Arid Environments 175, p.104082.
Sánchez-Montoya, M.M., Datry, T., Ruhi, A., Carlson, S.M., Corti, R., Tockner, K.
2023. Intermittent rivers and ephemeral streams are pivotal corridors for
aquatic and terrestrial animals. BioScience 73(4), 291-301.
Saltonstall, K., Lambert, A., Meyerson, L.A. 2010. Genetics and reproduction of
common (Phragmites australis) and giant reed (Arundo donax). Invasive
Plant Science and Management 3(4), 495-505.
Stefferud,
S.E., Reinthal, P.N. 2005. Fishes of
Aravaipa Creek, Graham and Pinal Counties, Arizona. Literature review and
history of research and monitoring. Report to U.S. Bureau of Land
Management, Safford, AZ. University of Arizona. Tucson, AZ. USA.
Stefferud,
S.E., Stefferud, J.A. 2007. Fish movement through intermittent stream
channels a case history study. Report to U.S. Bureau of Reclamation,
Phoenix, AZ. USA. Available at: https://www.usbr.gov/lc/phoenix/biology/azfish/pdf/intermittentStreams.pdf
Steward, A.L., Langhans, S.D., Corti, R., Datry, T. 2017. The biota of
intermittent rivers and ephemeral streams: Terrestrial and semiaquatic
invertebrates. In: Datry, T.,
Bonada, N., Boulton, A.J. (Eds.). Intermittent
Rivers and Ephemeral Streams, pp. 245-271. Academic Press.
Tolonen, K.E., Picazo, F., Vilmi, A., Datry, T., Stubbington, R., Pařil,
P., Perez Rocha, M., et al. 2019. Parallels and
contrasts between intermittently freezing and drying streams: from individual
adaptations to biodiversity variation. Freshwater Biology 64, 1679–1691
Van der Waal, B.C.W. 1997. Some observations on the fish
life in a seasonal sand river. Southern African Journal of Aquatic. Science 23:95–102
van Wilgen, N.J., Faulkner, K.T.,
Robinson, T.B., South, J., Beckett, H., Janion-Scheepers, C., Measey, J., et
al. 2022.
Climate Change and Biological Invasions in South Africa. In: Ziska, L.H.,
(Ed.). Invasive Species and Global Climate Change, 2nd
ed., pp. 158-187. CAB
International. Oxfordshire, UK.
Weyl, O.L.,
Ellender, B.R., Wassermann, R.J., Truter, M., Dalu, T., Zengeya, T.A. Smit,
N.J., 2020. Alien freshwater fauna in South Africa.
In: van Wilgen, B.W., Measey, J., Richardson, D.M., Wilson, J.R., Zengeya, T.A.
(Eds.). Biological Invasions in South Africa, pp. 153-183. Springer,
Berlin, Germany.
Williams, D.D. 2006. The biology of temporary waters. Oxford University Press.
348 pp.
Woodford, D.J.,
Hui, C., Richardson, D.M., Weyl, O.L. 2013.
Propagule pressure drives establishment of introduced freshwater fish:
quantitative evidence from an irrigation network. Ecological Applications
23(8), 1926-1937.
Zedler, J.B.,
Kercher, S. 2004. Causes and consequences of
invasive plants in wetlands: opportunities, opportunists, and outcomes. Critical
Reviews in Plant sciences 23(5), 431-452.