Introducción
Ante la situación de emergencia climática (Ripple et al. 2021) y crisis de biodiversidad sin
precedentes (Román-Palacios y Wiens 2020)
en la que nos encontramos, los científicos se enfrentan al reto de predecir y
anticipar con precisión el impacto del cambio climático en la biodiversidad.
Para cumplir con este gran desafío, varias estrategias y planes de acción
globales han delineado los objetivos de biodiversidad y cambio climático (IPBES
2019; United Nations 2019; Arneth et al. 2020). Sin embargo, estas estrategias
globales muestran ciertos sesgos e importantes omisiones entre sus objetivos.
Uno de los ejemplos más evidentes lo encontramos en los medios subterráneos, ya
que, siendo probablemente el ambiente no marino más extendido en la Tierra, han
sido sistemáticamente olvidados en estas estrategias globales (Sánchez-Fernández et al. 2021).
Para enfatizar la importancia y urgencia de proteger la
biodiversidad subterránea, el año 2021 fue elegido Año Internacional de las
Cuevas y el Karst. Con los ecosistemas subterráneos en el punto de mira, varios
científicos han realizado llamadas de atención lamentando su falta de
protección (ej. Mammola et al. 2019a),
mostrando que solo el 6.9% de los ecosistemas subterráneos conocidos se
encuentran bajo áreas protegidas en la superficie a nivel mundial (Sánchez-Fernández et al. 2021). Para
rectificar esta omisión, Wynne et al. (2021)
propusieron una hoja de ruta para guiar la conservación del bioma subterráneo
en los próximos años.
El medio subterráneo es uno de los ecosistemas menos
estudiados de la Tierra y solo recientemente estamos empezando a entender su
importancia eco-evolutiva e incluso social y económica (Mammola et al. 2020). Los ecosistemas subterráneos
brindan servicios ecosistémicos clave para la humanidad, como, por ejemplo, el
secuestro de gases de efecto invernadero, su uso potencial en la industria
farmacéutica y la ingeniería (Griebler y
Avramov 2015; Mammola et al. 2019a) o el
suministro de agua dulce (Danielopol et al. 2003).
De hecho, las aguas subterráneas representan el 95 % del agua dulce líquida
disponible (que no está acumulada en forma de casquetes polares y glaciares).
Se estima que alrededor del 50% del suministro actual de agua potable del mundo
proviene de aguas subterráneas, y que más de una cuarta parte de la población
humana depende total o parcialmente de esa agua potable (Giordano 2009). Además, más del 40% del agua dulce de
riego consuntiva total proviene de aguas subterráneas (Siebert et al. 2010). El medio subterráneo alberga
a especies de murciélagos que proporcionan servicios vitales (incluida la
dispersión de semillas, la polinización y la reducción de plagas agrícolas) (Kunz et al. 2011). Dentro de estos ecosistemas podemos
encontrar también una gran variedad de organismos especializados y singulares
que son a menudo endemismos con rangos de distribución muy restringidos, y
algunas especies representan ancestros de faunas que desaparecieron de los
hábitats superficiales (Culver y Pipan 2019).
Por lo tanto, no sorprende que los biólogos hayan estado fascinados durante
mucho tiempo por las peculiaridades de los organismos subterráneos (Darwin 1859; Jeannel 1943),
ya que muestran adaptaciones morfológicas y fisiológicas extremas que resultan
de gran valor para estudiar procesos evolutivos (Juan
et al. 2010). Por ejemplo, muchos de ellos han evolucionado hacia formas
ciegas, sin alas, sin pigmentos, etc., como consecuencia de severas
restricciones ambientales (Galán 2010). Además, dada
la extremada estabilidad climática del medio subterráneo profundo, donde tanto
la humedad (próxima al punto de saturación), como la temperatura, varían muy
poco a lo largo del día y del año (Badino 2010; Culver y Pipan 2019), las especies subterráneas
son generalmente estenohigrobias (es decir, muy sensibles a la desecación) y
estenotermas (es decir, presentan rangos de tolerancia térmica mucho más
estrechos que las especies de la superficie) (Howarth
1980; Hadley et al. 1981; Howarth y Moldovan 2018).
La fauna subterránea es un componente clave de la
biodiversidad global, dado su alto grado de endemismo y de especialización, y
su singularidad filogenética y funcional. Sin embargo, la biodiversidad
subterránea se encuentra actualmente amenazada por diversas actividades
humanas, y puede ser especialmente sensible al cambio climático (Mammola et al. 2019a). De hecho, aunque se
trata de sistemas en general muy aislados e inaccesibles, el medio subterráneo
no está desvinculado de los impactos que afectan a otros ecosistemas en la
superficie ni de los efectos del cambio climático. Precisamente por el alto
grado de especialización de su fauna, ante un aumento rápido de la temperatura
y un cambio en las condiciones climáticas, los hábitats subterráneos pueden
convertirse en trampas sin salida para algunas de las especies que actualmente
viven allí de forma obligada, pero también pueden actuar como refugios para
algunas especies de la superficie, lo que daría lugar a importantes
modificaciones de las interacciones bióticas en el subsuelo (Mammola et al. 2019b;
Nicolosi et al. 2023).
La península ibérica es una zona de gran interés con
respecto a su fauna hipogea o cavernícola (Culver
y Pipan 2019; Reboleira et al. 2011; Iannella et al. 2020). De hecho, Salgado et al. (2022) han elaborado por primera vez
un listado completo de taxones descritos hasta la fecha en el área
ibero-balear, que incluye un total de 1285 especies y 79 subespecies hipogeas.
Ante los indicios que muestran que las especies subterráneas podrían ser particularmente
sensibles al cambio climático (Mamola et al. 2019b),
el interés científico por este medio ha aumentado de forma considerable, 'desarrollándose
en los últimos años un buen número de estudios destinados a conocer la
capacidad de respuesta y anticipar los efectos del cambio climático en la
biodiversidad subterránea (Vaccarelli et al. 2023).
Para tomar decisiones efectivas, los gestores del medio natural necesitarán
modelos y predicciones tan precisos como sea posible, así como poder acceder de
forma sintética a toda la información científica que se está generando en la
actualidad (Grimm et al. 2020; Mammola et al. 2022).
En este artículo proporcionamos una revisión sistemática de
los estudios que exploran los efectos del cambio climático en la biodiversidad
subterránea ibérica, para (i) describir su variabilidad en cuanto a enfoques
experimentales, ambientes estudiados, niveles de organización biológica, grado
de especificidad por el medio subterráneo y respuestas estudiadas y (ii)
determinar otros factores que pueden interactuar con un aumento de la
temperatura en los ecosistemas subterráneos. Al sintetizar el conocimiento
actual sobre los efectos del cambio climático en los organismos subterráneos de
la península ibérica, podremos hacer un primer balance de los progresos
realizados, identificar vacíos de conocimiento, así como destacar las
necesidades de investigación en ecología subterránea, proporcionando
información dirigida a mejorar el seguimiento, la gestión y la restauración de
los ecosistemas subterráneos ibéricos.
Materiales y métodos
Búsqueda bibliográfica sistemática y criterios de
inclusión de artículos
En septiembre de 2022 se realizó una búsqueda bibliográfica
estandarizada en la base de datos Web of Science, incluyendo grupos de
términos relativos a i) los tipos de ecosistemas subterráneos, ii) organismos
subterráneos y su especificidad por este medio, iii) área de estudio, iv)
cambio climático (variables ambientales y tipo de respuestas biológicas frente
al cambio climático). Tras varias pruebas, llegamos a un acuerdo con la
siguiente cadena de búsqueda:
TS =
(“MSS” OR “Shallow subterranean habitat” OR “Superficial subterranean habitat”
OR “aquifer*” OR “cave*” OR "groundwater*" OR
"subterranean" OR “underground” OR “roost”)
AND TS
= (“species” OR “endemic” OR “obligate” OR “facultative” OR "stygo*"
OR "troglo*” OR “eutroglo*” OR “eustygo*”)
AND CU
= (“Iberian” OR “Pyrenees” OR “Spain” OR “Spanish” OR “Portugal”)
AND TS
= ("climate change" OR "global warming" OR “climate
warming” OR “temperature increase” OR “temperature rise” OR “relative humidity
change” OR “salinity increase” OR “salinity change” OR “relative humidity
decrease” OR “climatic variability hypothesis”)
AND TS
= (“thermal tolerance” OR “thermal limits” OR “CTmax” OR “CTmin” OR “LTmax” OR
“LTmin” OR “thermal stress” OR “thermal shock” OR “acclimat*” OR “physiol*” OR
“ecophysiolog*” OR “survival” OR “mortality” OR “fitness” OR “metaboli*” OR
“locomot*” OR “growth” OR “development*” OR “size” OR “gene expression” OR
“genetic” OR “enzym*” OR “proteomic” OR “hormonal” OR “oxidative” OR
“behavior*” OR “behaviour*” OR “reproduct*” OR “life-cycle” OR “immune” OR
“distribution*” OR “dispersal” OR “range” OR “population” OR “community” OR
“species composition” OR “occurrence” OR “habitat*” OR “extinction” OR
“decline” OR “migration” OR “refugium” OR “refugia” OR “shelter” OR “abundance”
OR “presence” OR “richness” OR “competition” OR “interaction” OR “phenolog*” OR
“season*” OR “circadian” OR “heat shock” OR “heat-shock” OR “adaptation” OR
“drift” OR “alien species” OR “invasion” OR “species turnover” OR “thermophile”
OR “stenothermal” OR “warm-dwelling” OR “sensitivity” OR “thermal niche” OR
“pathogen*” OR “parasite*” OR “virus*” OR “viral”).
Se examinaron todos los títulos y resúmenes de los artículos
resultantes (y el cuerpo del artículo cuando era necesario consultar más
detalles), incluyéndose aquellos que
cumplían los siguientes criterios:
i) están
centrados en especies subterráneas (obligadas o facultativas) de cualquier
grupo taxonómico,
ii) estudian los
efectos del cambio climático sobre los organismos subterráneos, considerando
los cambios ambientales asociados a este fenómeno que pueden afectar al medio
subterráneo (aumento de la temperatura o cambios en el régimen de variabilidad
térmica, en la humedad relativa, o cambios en la salinidad en ambientes
acuáticos subterráneos) e incluyendo diversos tipos de respuestas a distintos
niveles de organización, desde el individuo a la comunidad: respuestas
fisiológicas (ej. límites térmicos fisiológicos, capacidad de aclimatación
térmica, tasa metabólica), de comportamiento (ej. actividad locomotora),
moleculares (respuestas hormonales, enzimáticas, proteómicas o genómicas),
respuestas relacionadas con la reproducción, el crecimiento, ciclo de vida,
fenología, cambios en el rango de distribución, dispersión, cambios en la
estructura y composición de las comunidades, declives poblacionales,
migraciones, extinciones, interacciones bióticas (ej. especies invasoras,
patógenos),
iii) el área de
estudio es parte o la totalidad de la península ibérica o, en el caso de
artículos que abarcan áreas de estudio más extensas, proporcionan información
sobre los efectos del cambio climático en especies, poblaciones o comunidades
subterráneas ibéricas.
Además de los
artículos que no cumplían los anteriores criterios, se excluyeron aquellos que
no aportaban información suficiente para extraer conclusiones sobre el efecto
del cambio climático, cuantitativa o cualitativamente, en el organismo(s) y
respuesta(s) concreta estudiados.
Búsqueda bibliográfica adicional
Puesto que la
literatura sobre los efectos del cambio climático en el medio subterráneo es
escasa, en particular en la península ibérica, se realizaron varias búsquedas
bibliográficas adicionales, no estandarizadas. En primer lugar, se hicieron
varias búsquedas en Google Scholar específicas para estudios sobre
murciélagos cavernícolas, ya que los artículos de este tipo encontrados en la
búsqueda estandarizada fueron escasos. En segundo lugar, se revisó la
bibliografía citada en los artículos que cumplían los criterios de inclusión.
Extracción de datos
De cada artículo se extrajo la siguiente información: área
de estudio, clasificación taxonómica y ecología de los taxones estudiados,
metodología aplicada (tipo de método, variables independientes y variables
respuesta analizadas, rango de temperaturas testadas en el caso de estudios
experimentales, incluyendo la temperatura del tratamiento control, en su caso,
y la temperatura máxima) y resultados cuantitativos (en los artículos que
aportaban datos suficientes). Dada la diversidad de metodologías aplicadas, se
intentó presentar los resultados de manera comparable, al menos entre casos de
estudio que emplearon el mismo tipo de aproximación metodológica. Por ejemplo,
para casos de estudio donde se estimaron límites térmicos letales, se extrajo
directamente el valor de los mismos, como indicador representativo de la
sensibilidad de las especies al aumento de la temperatura. Para casos de
estudio en los que se midieron experimentalmente otro tipo de respuestas tras
un periodo de aclimatación a distintas temperaturas, extrajimos el valor de la
variable respuesta correspondiente en el tratamiento control y en la
temperatura máxima testada, y calculamos la tasa de cambio porcentual de la
variable respuesta con relación al cambio de temperatura entre dichos
tratamientos. En este caso, un valor negativo indica una disminución de la
variable respuesta al aumentar la temperatura (indicando generalmente un efecto
de la temperatura perjudicial para el organismo), y un valor positivo indica un
aumento (reflejando generalmente una capacidad de ajuste por aclimatación). En
estudios que estimaron cambios en el rango de distribución de las especies en
escenarios de cambio climático utilizando modelos de distribución de especies,
se extrajo, cuando fue posible, el porcentaje de área favorable que se perderá
o ganará. En estos casos, cuando se presentaban predicciones para distintos
escenarios de cambio climático, por simplificación, se escogieron las
predicciones para el periodo temporal más lejano y el escenario futuro de cambio
climático correspondiente al contexto socioeconómico y trayectoria de emisión
de gases de efecto invernadero del IPCC más pesimista, puesto que los informes
más recientes del IPCC indican que el ritmo actual de emisiones ya supera
dichos escenarios (IPCC 2022).
Resultados
Resultados de la búsqueda y cribado de artículos
El número de artículos obtenidos de la búsqueda
bibliográfica en Web of Science y de la búsqueda adicional fue de 135 y 14 artículos, respectivamente. De éstos,
sólo 15 (un 10%) fueron seleccionados tras descartar los que no cumplían los
criterios de inclusión establecidos (listado disponible en Material
Suplementario). Todos los estudios revisados han
sido publicados en los últimos 10 años (de 2013 en adelante) (Fig. 1A).
En la Tabla
A1 del anexo se muestra un resumen de la
información recopilada en los artículos revisados (todos los datos recopilados
están disponibles en Material
Suplementario).
Organismos y hábitats
El conjunto de datos analizado incluye un total de 135
taxones. Todos los estudios revisados han utilizado animales como modelo de
estudio, restringiéndose a dos grupos concretos: murciélagos entre los
vertebrados (11 especies) y principalmente
coleópteros (también algunos arácnidos, colémbolos y diplópodos) entre los
invertebrados (124 especies) (Tabla A1, Fig. 1B).
Todas las especies estudiadas son terrestres, con distintos
grados de especialización al medio subterráneo (Fig. 1C),
destacando la ausencia de estudios con especies acuáticas subterráneas.
Tipos de respuestas estudiadas
Los artículos revisados se pueden agrupar en dos grupos
claramente diferenciados en cuanto al tipo de respuesta estudiada: artículos
basados en experimentos fisiológicos que exploran aspectos relacionados con la
tolerancia térmica de las especies subterráneas, como medida de la capacidad de
estas para hacer frente al cambio climático dentro de su rango actual de
distribución (capacidad de persistencia), y artículos que estudian cambios en
el rango de distribución en distintos escenarios simulados de cambio climático
basados en el modelado de nicho ecológico (Fig. 1D).
Del primer grupo de artículos se han recopilado 42 casos de estudio en los que
se han utilizado exclusivamente invertebrados (mayoritariamente coleópteros)
como modelo de estudio, mientras que los estudios sobre cambios corológicos han
estado centrados principalmente en murciélagos (15
casos de estudio), excepto algunos que han utilizado invertebrados (4 casos de
estudio) (Tabla A1). Ningún artículo ha
analizado respuestas comportamentales, reproductivas, del ciclo de vida o
fenológicas en especies o poblaciones subterráneas ibéricas. En nuestra
revisión tampoco se han encontrado estudios a nivel de comunidad, ni relativos
a interacciones biológicas (ej. especies invasoras o patógenos), en relación
con el cambio climático.
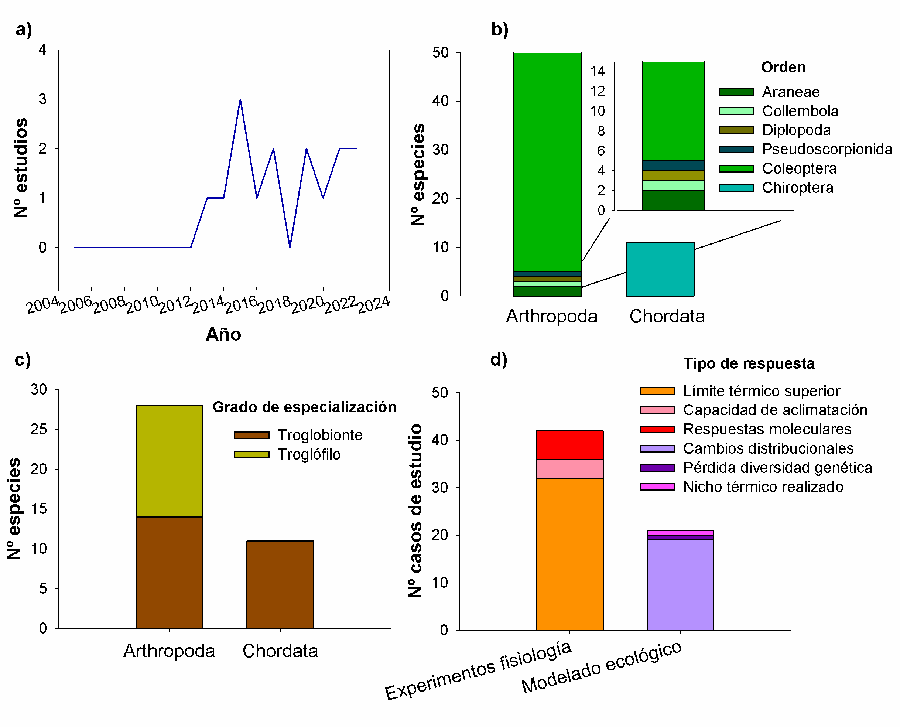
Figura 1. Descripción general de los estudios
sobre los efectos del cambio climático en organismos subterráneos en la
península ibérica: A) número de estudios publicado por año, B)
grupo taxonómico, C) grado de especialización al medio subterráneo y D)
tipo de respuesta estudiada.
Figure 1. General description of the studies on the effects of climate
change on subterranean organisms in the Iberian Peninsula: A) number of studies published per year,
B) taxonomic group, C) degree of specialization to the
subterranean environment and D) studied response type.
A continuación, se describen con más detalle los estudios
correspondientes a los dos grupos de respuestas principales:
Tolerancia térmica (capacidad de persistencia)
De la revisión bibliográfica se han extraído siete artículos en cuya metodología se han
llevado a cabo experimentos de laboratorio con el objetivo de estudiar la
capacidad fisiológica de las especies subterráneas de hacer frente al cambio
climático. Todos estos estudios se han centrado en invertebrados subterráneos
(22 especies), principalmente coleópteros, pero también colémbolos, arácnidos y
miriápodos, y se han estudiado diferentes tipos de respuestas fisiológicas al
aumento de temperatura (Tabla A1). No se han
considerado otros factores aparte de la temperatura.
En seis artículos se ha
analizado la supervivencia de especies subterráneas expuestas a diferentes
temperaturas, estimando: i) límite térmico superior letal, utilizando
generalmente la LT50 (temperatura letal para el 50% de los
individuos expuestos) como variable respuesta; y/o ii) capacidad de
aclimatación térmica (capacidad de aumentar el límite superior letal o el
tiempo de supervivencia durante una exposición a estrés térmico tras un periodo
de aclimatación) (Rizzo
et al. 2015; Pallarés et al. 2019, 2020, 2021; Colado et al. 2022a, 2022b). De
manera general, estos estudios han mostrado que las especies más especializadas
al medio subterráneo (es decir, troglobiontes o subterráneos obligados, y entre
éstos, los que viven en las partes más profundas de las cavidades) tienen una
menor tolerancia térmica (LT50 con valores entre 15-20ºC) que las
especies troglófilas (subterráneas facultativas) con un grado de
especialización menor (LT50 por encima de los 21ºC) (Tabla A1). En cuanto a la capacidad de
aclimatación, Pallarés et al. (2021) mostraron
que varias especies subterráneas fueron capaces de aumentar, aunque muy
ligeramente, su límite térmico superior tras una aclimatación a una temperatura
alta, siendo la capacidad de aclimatación mayor en las especies con menor grado
de especialización subterránea (Tabla A1). En
estudios donde se midió la capacidad de aumentar el tiempo de supervivencia
durante un estrés térmico tras una exposición previa a distintas temperaturas,
varias especies subterráneas (varios coleópteros y un diplópodo) mostraron una
respuesta de aclimatación positiva (el tiempo de supervivencia aumentó varios
días en los individuos previamente aclimatados a temperaturas más altas) (Rizzo et al. 2015; Pallarés
et al. 2019, 2020). Sólo en una de las
especies estudiadas (el colémbolo Deuteraphorura silvaria), el tiempo de
supervivencia disminuyó tras la aclimatación a una temperatura alta (Pallarés et al. 2019) (Tabla
A1).
Tan solo uno de los artículos analizados (Pallarés et al. 2020) incluyó otro tipo de
respuestas, de tipo molecular, además de la estimación de límites térmicos
letales y capacidad de aclimatación. Este estudio analizó la respuesta a un
aumento de la temperatura de cinco biomarcadores de estrés oxidativo y de la
acetilcolinesterasa (enzima asociada a la neurotoxicidad) en el coleóptero leiódido
Parvospeonomus canyellesi. Los autores mostraron que la exposición a una
temperatura no letal indujo cambios significativos en la actividad de las
enzimas glutation S-transferasa y acetilcolinesterasa, así como en la capacidad
antioxidante total y el glutatión (Tabla A1), que
se interpretaron como efectos subletales (estrés oxidativo y nervioso), aunque
la especie fue capaz de desarrollar una respuesta antioxidante.
Cambios en el rango de distribución
Un total de 10 artículos
han aplicado modelos de distribución de especies en distintos escenarios de
cambio climático para predecir cambios en el área climáticamente favorable. La
mayoría de estos estudios (8) se han
centrado en murciélagos, proporcionando datos para 11 especies. En general,
estos estudios predicen una contracción del rango de distribución ibérico para
la mayoría de quirópteros estudiados (aunque en la mayoría de casos no se
aportan datos cuantitativos de la proporción de área favorable que se podría
perder; ver Tabla A1), y un desplazamiento de las
condiciones climáticamente favorables hacia el norte peninsular.
Amorim et al. (2014)
predijeron que una importante proporción del área climáticamente favorable para
tres especies de murciélagos (Barbastella barbastellus, Hypsugo savii,
Myotis daubentonii) en el norte de Portugal desaparecerá en 2080, mientras
que no habrá nuevas zonas favorables o estas serán muy reducidas en el caso de B.
barbastellus dentro del área de estudio. Un trabajo más reciente que abarcó
un área de estudio más extensa, basado en el rango de distribución europeo de
varias especies, también predice una importante contracción del rango de
distribución ibérico de M. daubentonii, especialmente en el norte de la
península, en 2070, al igual que para Plecotus auritus (McGowan et al. 2021). Ese mismo estudio arroja
predicciones menos dramáticas para Pipistrellus pipistrellus y Rhinolophus
hipposideros, aunque anteriormente Nagy et al. (2017)
estimaron una importante pérdida de área favorable para P. pipistrellus en
el sur, sureste y suroeste de la península ibérica para el mismo periodo.
Sólo tres de los artículos revisados han proporcionado
estimas cuantitativas de la proporción de área climáticamente favorable que
perderán o ganarán especies de murciélagos en distintos escenarios de cambio
climático. Razgour
(2015) estimó que como
resultado de la pérdida de hábitat favorable para Plecotus austriacus,
un 64% de las colonias actuales de esta especie en la península ibérica
se encontrarán en condiciones climáticas desfavorables en 2070. En la misma
dirección, Razgour et al. (2015) estimaron que un 16% del área climáticamente favorable para Myotis
escalerai en la península ibérica se volverá desfavorable en 2070 (la mayor
parte de su rango de distribución actual en el interior y el sur de la
península), mientras que zonas en las que la especie no está actualmente
presente (la costa atlántica norte, Pirineos y noroeste de Francia) tendrán
condiciones favorables. Más recientemente, considerando las adaptaciones
climáticas locales de las especies en modelos de distribución que incorporan la
variación intraespecífica (es decir, diferenciando entre genotipos adaptados a
clima árido y cálido o frío y húmedo), Razgour et al. (2019) proyectaron que un 19 y un
58% del rango de distribución ibérico de M. escalerai y M. crypticus,
respectivamente, podría perderse en 2070 (Tabla A1).
En la mayoría de los casos
descritos anteriormente, las predicciones de cambios en el rango de
distribución de las especies se han basado en el modelado de su nicho ecológico
realizado (es decir, se ha estimado el área que será favorable para la especie a
partir de las condiciones ambientales ocupadas), sin tener en cuenta otros
aspectos que también condicionan la distribución de las especies. Como
excepción, Razgour et al. (2013) consideraron la capacidad de adaptación local de la
especie estudiada (Plecotus austriacus) en
sus predicciones, incorporando la diversidad genética en los modelos de
distribución. Estos modelos predijeron una pérdida del área
climáticamente favorable para esta especie en la península ibérica en 2080, que
además estaría asociada a una importante pérdida de diversidad genética
(estimada como el número de haplotipos únicos
que se encontrarán en zonas desfavorables en el futuro).
Tres de los
artículos revisados aplicaron la metodología del modelado ecológico para
predecir cambios en el rango de distribución de especies de invertebrados
subterráneos. Mammola e Isaia (2017) modelaron la distribución de dos arañas cavernícolas (Meta
bourneti y M. menardi), teniendo en cuenta la competencia potencial
entre las dos especies. Los modelos predijeron una importante regresión de las
poblaciones exclusivas de cada especie en la península
ibérica, y también las áreas
donde coexisten, en 2070. Sánchez-Fernández
et al. (2016) modelaron la
distribución presente y futura de 83 especies de un clado de escarabajos de la
familia Leiodidae, y estimaron que casi un 70% de estas especies no mantendrán sus poblaciones en condiciones
climáticamente favorables en 2080 (Tabla A1). Colado
et al. (2022a) estimaron que, en
2070, el riesgo de extinción local sería alto para al menos la mitad de las
poblaciones del pseudoescorpión subterráneo Neobisium vasconicum
teniendo en cuenta el nicho térmico realizado de esta especie, cuyo margen
térmico de seguridad sería muy estrecho (menos de 1ºC) de acuerdo con los datos
climáticos en su área actual de distribución. Sin embargo, Sánchez-Fernández et al. (2016) y Colado
et al. (2022a) también estimaron
experimentalmente la tolerancia térmica de estas especies, y sus resultados
indican que serían capaces de vivir a temperaturas superiores que las estimadas
modelando los nichos realizados, lo cual en principio les permitiría hacer
frente al aumento de temperatura en el futuro (ver apartado anterior: Tolerancia térmica
(capacidad de persistencia)).
Discusión
A pesar de la
importancia de estudiar los efectos del cambio climático en el medio
subterráneo y de las ventajas que este medio ofrece para ello (Sánchez-Fernández
et al. 2018; Mammola et al. 2019b), este trabajo muestra que en la península ibérica se le ha empezado a prestar atención a este tema en los
últimos 12 años. Además, esta revisión localiza importantes carencias en
nuestro conocimiento sobre los efectos del cambio climático en la biodiversidad
subterránea ibérica y propone una serie de líneas prioritarias en las que se
debería trabajar en los próximos años.
¿Qué se sabe
sobre los efectos del cambio climático en la biodiversidad subterránea ibérica?
Muy pocos
estudios han tratado de predecir los efectos del cambio climático en los
ecosistemas subterráneos ibéricos. Además de la escasez generalizada de
estudios en este campo, existen importantes sesgos entre los realizados. En
primer lugar, existe un sesgo hacia dos tipos de aproximaciones metodológicas
centradas en dos tipos de respuestas concretas, y dentro de cada respuesta,
también se puede apreciar un sesgo en el grupo taxonómico estudiado. Los
estudios de tipo experimental se han centrado en la fisiología térmica y se han
realizado exclusivamente con invertebrados, mientras que los estudios en los
que se evalúa el cambio potencial del área de distribución se han realizado
principalmente con murciélagos. La ausencia de estudios fisiológicos con
vertebrados es con seguridad consecuencia de las dificultades para realizar
estudios experimentales con estos organismos, donde los diferentes tratamientos
requieren de un número mínimo de individuos que es más fácilmente abordable
trabajando con invertebrados. Además, los estudios experimentales con
vertebrados requieren el uso de técnicas no invasivas, que seguramente pueden
ser más complejas de implementar.
Los estudios
experimentales revisados han analizado casi exclusivamente la tolerancia
térmica de las especies, un rasgo clave para estimar su capacidad para hacer
frente al cambio climático dentro de su rango actual de distribución (capacidad
de persistencia) (Arribas et al. 2012,
2017). En general, estos estudios sugieren que
las especies subterráneas son capaces de tolerar temperaturas bastante
superiores a las que están expuestas en su medio. De acuerdo con estos
resultados, la mayoría de las especies estudiadas tendrían la capacidad
fisiológica de hacer frente al aumento de temperatura en su hábitat. Sin
embargo, como ocurre en general entre las especies más especialistas (Clavel et al. 2011), esa capacidad se reduce en las especies
con mayor grado de especificidad por el medio subterráneo (subterráneas
obligadas y dentro de estas, las que ocupan las partes más profundas de las
cuevas) (ej. Colado
et al. 2022b). Esta
asociación entre la tolerancia térmica y el grado de especialización al medio
subterráneo (relacionado con la variabilidad térmica que experimentan las
especies) también ha sido demostrada en otros linajes de invertebrados
subterráneos fuera de la península ibérica (Raschmanová et al. 2018, Mammola et al. 2019c). Sin embargo, estas conclusiones se basan
exclusivamente en el estudio de dos variables concretas: la supervivencia
(límites térmicos letales) y capacidad de aclimatación térmica, que se han
medido generalmente en periodos de exposición relativamente cortos (desde días
a semanas). El único estudio que ha explorado respuestas subletales (estrés
oxidativo) en una especie subterránea sugiere que un aumento de la temperatura,
dentro del rango de tolerancia de la especie, podría tener efectos fisiológicos
negativos (Pallarés
et al. 2020). Por lo
tanto, es fundamental ampliar el estudio de este tipo de respuestas subletales
para obtener estimas más realistas de los rangos de temperatura que podrían ser
fisiológicamente más favorables para las especies subterráneas. Así mismo,
tampoco se han explorado otros componentes del fitness (ej. crecimiento,
reproducción), ni respuestas a nivel genómico, o los efectos del aumento de
temperatura a largo plazo. Uno de los principales retos para cubrir esta laguna
de conocimiento es el de adaptar este tipo de aproximaciones experimentales a
las especies subterráneas, difíciles de capturar en su ambiente, con ciclos de
vida relativamente largos y cuya cría en laboratorio resulta muy compleja. En
este sentido, será clave el desarrollo de metodologías que permitan obtener
resultados robustos trabajando con tamaños de muestra pequeños.
Por otro lado, el
uso de modelos de distribución de especies prevé importantes reducciones
generalizadas en las áreas de distribución futura de murciélagos que anidan en
cuevas en la península ibérica. Este tipo de metodología presenta
ciertas ventajas cuando se trabaja en el medio subterráneo, especialmente para
especies troglobiontes (subterráneas obligadas), ya que, por ejemplo, se
minimiza la incertidumbre de la influencia de las interacciones bióticas puesto
que las comunidades son muy sencillas, y se conocen, de manera más precisa que
en la superficie, las condiciones ambientales a las que están expuestos los
organismos (Mammola
y Leroy 2018). Sin
embargo, estos modelos también presentan algunas limitaciones para especies
subterráneas con baja capacidad de dispersión (ej. invertebrados). En primer
lugar, la falta de conocimiento preciso de las distribuciones de las especies
puede condicionar las estimas del nicho ecológico que se derivan de esos datos
distribucionales parciales (Jiménez-Valverde et al. 2008; Peterson et al. 2011). Por otro lado, estos modelos podrían sobrestimar la
cantidad de área con clima favorable tanto en el presente como en el futuro, ya
que una buena cantidad de estas zonas nunca serán accesibles debido a las
limitaciones de dispersión, es decir, en estas especies probablemente se
maximiza la diferencia entre distribución potencial y real. Atendiendo a las
limitaciones de dispersión de la fauna subterránea obligada, más que localizar
nuevas áreas con clima favorable en el futuro, requiere especial atención conocer
la capacidad de persistencia de las especies en las localidades actuales, y
para ello, el uso combinado de modelos de distribución de especies y
experimentos de fisiología resultará de especial importancia.
Vacíos de
conocimiento en el estudio de las respuestas de la biodiversidad subterránea ibérica
al cambio climático y perspectivas futuras
Además de los
sesgos anteriormente descritos, en esta revisión se han detectado importantes
omisiones en el estudio de los efectos del cambio climático sobre la
biodiversidad subterránea. Por ejemplo, en los estudios experimentales sólo se
ha evaluado el efecto del aumento de la temperatura. La valiosa información
proporcionada por los estudios de tolerancia térmica podría refinarse aún más
teniendo en cuenta otros factores, como humedad, salinidad, contaminantes,
etc., y sus posibles interacciones con un aumento de temperatura. Especialmente
relevante podría ser el estudio del efecto de la humedad, cuyos altos valores
en el medio subterráneo (en el punto de saturación o muy próximo al mismo)
parecen ser esenciales para la supervivencia de los organismos troglobiontes (Mammola et al. 2019b). Howarth (1980) sugirió que los invertebrados subterráneos tienen una alta
permeabilidad cuticular, asociada con una resistencia muy baja a la desecación,
lo cual ha sido demostrado empíricamente en algunas especies (Ahearn y Howarth 1982; Yoder et al. 2011). Estudios experimentales sobre la importancia de un
descenso en la humedad relativa, así como el efecto de su interacción con
un aumento de temperatura en la supervivencia de las especies
subterráneas, proporcionarán información de gran utilidad para comprender mejor
el impacto del cambio climático sobre la biodiversidad subterránea.
La mayoría de las
especies estudiadas (sobre todo en experimentos de fisiología) se encuentran en
el norte de la Península, en las zonas de los Pirineos y la Cordillera
Cantábrica. Si bien el estudio de estas regiones es clave en un contexto
ibérico, desconocemos si las respuestas de la fauna subterránea del área
meridional (sierras béticas-sistema ibérico) son similares a las del norte. Es
especialmente interesante realizar un seguimiento a las especies que viven en
cuevas que se encuentran en temperaturas próximas o superiores a los 20ºC, ya
que los límites térmicos estimados para muchas especies del Pirineo están
próximos a esa cifra, y el rango térmico de seguridad en las especies del sur
podría ser muy pequeño, por lo que sutiles cambios en las condiciones
climáticas de las cuevas podrían provocar efectos fisiológicos importantes,
reduciendo sus poblaciones.
La mayor parte de
los estudios revisados se han realizado a nivel de especie, y solo unos pocos a
nivel de población, mientras que no existe información sobre respuestas a nivel
de comunidad. El uso de modelos jerárquicos de comunidades (Hierachical
Modelling of Species Communities - HMSC), podría permitir la integración de
datos de ecología de comunidades con datos ambientales, rasgos de las especies,
proporcionando así información predictiva sobre los procesos de ensamblaje de
las comunidades (Tikhonov et al. 2020).
Esto es relevante, ya que las interacciones bióticas (competencia, mutualismo)
pueden tener una importante influencia en la supervivencia de especies
subterráneas ibéricas (ver Mammola e Isaia 2017) y estas interacciones pueden verse afectadas por los
cambios ambientales. Este tema cobra especial relevancia si tenemos en cuenta
el proceso de colonización del medio subterráneo por parte de especies
típicamente superficiales como refugio ante el cambio climático. De hecho, la
presencia de especies de la superficie en ambientes subterráneos ya ha sido
documentada en algunos lugares, por ejemplo, la rana ibérica en Serra da
Estrela, Portugal (Rosa y Penado 2013).
Tampoco se tienen
datos sobre la importancia de los patógenos en el medio subterráneo y si el
cambio climático intensificará o atenuará su importancia en el contexto
ibérico, aunque estudios en otras regiones han mostrado la influencia del clima
en la expansión de patógenos en estos ambientes (ej. Escobar et al. 2014). Los organismos subterráneos podrían ser
muy susceptibles a los patógenos, ya que su capacidad inmune puede estar
reducida, por varios motivos: i) por el elevado gasto energético que suponen
las respuestas inmunológicas en un medio con recursos limitados, ii) por una
presión selectiva débil, ya que al vivir en comunidades simplificadas podrían
estar expuestos a una menor presión patogénica que las especies de la
superficie, y iii) en el caso de los invertebrados subterráneos, estos tienen
además cutículas muy finas que podrían ser menos eficientes como barrera frente
a la entrada de patógenos. Por lo tanto, la entrada de patógenos directa (ej. a
través de agua contaminada) o indirecta (por medio de otras especies invasoras
que introducen sus propios patógenos) puede ser una importante amenaza para la
fauna subterránea.
Existe un
importante desconocimiento sobre los efectos del cambio climático en la fauna
subterránea acuática, ya que todas las especies ibéricas estudiadas son
terrestres. Aun así, se espera que la vulnerabilidad de la biodiversidad
acuática subterránea sea alta, debido no sólo al cambio climático, sino a una
combinación de factores derivados de la sobreexplotación y contaminación de los
acuíferos. Por tanto, las especies de las aguas subterráneas podrían ser muy
sensibles a los impactos antropogénicos y a los cambios ambientales,
convirtiéndose en importantes bioindicadores de la calidad de las aguas
subterráneas. Aumentar la información y el número de estudios sobre la fauna
subterránea acuática y su respuesta frente al cambio climático aportaría una
valiosa información a los gestores y los responsables de la toma de decisiones
sobre el estado de estos importantes ecosistemas (Griebler y Avramov
2015).
Conclusiones
El estudio de los
efectos del cambio climático en la biodiversidad subterránea ibérica se
encuentra en un estadio muy inicial, por lo que existen importantes lagunas y
sesgos de conocimiento sobre este tema. Los estudios revisados se han centrado
en la fisiología térmica en invertebrados, para estimar su capacidad
fisiológica de hacer frente a un aumento de temperatura, o en la predicción de
cambios en el área de distribución en distintos escenarios de cambio climático,
utilizando sobre todo murciélagos como modelo de estudio. Los esfuerzos de
investigación en este campo deberían dirigirse hacia: (i) ampliar el rango
taxonómico y geográfico de las especies estudiadas, utilizando el mejor modelo
de estudio para cada aproximación metodológica y objetivo de investigación,
incluyendo especies acuáticas subterráneas y poblaciones del centro y sur de la
península ibérica, (ii) explorar un mayor rango de
respuestas, incluyendo, por ejemplo, respuestas subletales y a largo plazo
frente al estrés térmico, efectos de la temperatura en el crecimiento,
comportamiento, reproducción, ciclo de vida y otros aspectos relacionados con
el fitness, respuestas a nivel de comunidad e interacciones biológicas,
(iii) considerar los efectos de otros factores (humedad, salinidad, contaminantes)
que pueden interaccionar con un aumento de la temperatura. Mejorar
nuestro conocimiento sobre los ecosistemas subterráneos es un primer paso
fundamental para proponer acciones de conservación basadas en evidencias
científicas (Mammola et al. 2022) y preservar
los importantes servicios ecosistémicos que nos brindan estos ambientes. Estas recomendaciones se deberían alinear
con una consideración de los ecosistemas subterráneos en las estrategias y
objetivos globales y nacionales de conservación de la biodiversidad y acción
frente al cambio climático.
Contribución de los autores
Raquel Colado,
David Sánchez Fernández, Susana Pallarés: Conceptualización, Investigación, Metodología, Redacción -
borrador original, Redacción - revisión y edición.
Agradecimientos
Este artículo es parte del proyecto de I+D+i
(PID2021-124640NB-I00), financiado por MCIN/ AEI/10.13039/501100011033/ “FEDER
Una manera de hacer Europa”. R.C está financiada por una beca predoctoral del
Ministerio de Ciencia e Innovación de España (FPI). D.S.-F. está financiado por
el programa Ramón y Cajal (RYC2019-027446-I) MCIN/AEI/ 10.13039/501100011033
“FSE Invierte en tu futuro” del Ministerio de Ciencia e Innovación de España.
S.P. está financiada por un contrato postdoctoral de la “Consejería de Economía,
Conocimiento, Empresas y Universidad de la Junta de Andalucía-Fondo Social
Europeo de Andalucía 2014-2020” (SP-DOC_01211). Agradecemos a Alberto
Sendra y Miguel López Munguira las constructivas revisiones realizadas a este
manuscrito, así como a Alberto Jiménez-Valverde y Vicente Ortuño por coordinar
y editar este número especial.
Referencias
Ahearn,
G.A., Howarth, F.G. 1982. Physiology
of cave arthropods in Hawaii. Journal of Experimental Zoology 222:
227-238.
Amorim, F., Carvalho, S.B., Honrado, J., Rebelo, H. 2014. Designing optimized multi-species monitoring networks to detect
range shifts driven by climate change: a case study with bats in the North of
Portugal. PloS one 9(1): e87291.
Arneth, A., Shin, Y.J., Leadley, P., Rondinini, C., Bukvareva, E., Kolb,
M., et al. 2020. Post-2020 biodiversity targets need to embrace climate change.
Proceedings of the National Academy of Sciences 117: 30882-30891.
Arribas, P., Abellán, P.,
Velasco, J., Bilton, D.T., Millán, A., Sánchez-Fernández, D. 2012. Evaluating drivers of vulnerability to climate change: a guide for
insect conservation strategies. Global Change Biology 18:
2135–2146.
Arribas, P., Abellán, P.,
Velasco, J., Millán, A., Sánchez-Fernández, D. 2017. Conservation
of insects in the face of global climate change. En: Johnson, S.N., Jones, T.H.
(Eds.), Global Climate Change and Terrestrial Invertebrates, pp. p.
349–367. John Wiley & Sons, Ltd, Chichester, Reino Unido.
Badino, G. 2010. Underground meteorology. What’s the weather underground? Acta
Carsologica 39: 427– 448.
Clavel, J., Julliard, R.,
Devictor, V. 2011. Worldwide decline of specialist species:
toward a global functional homogenization?. Frontiers in Ecology and the
Environment 9: 222-228.
Colado, R., García-Meseguer, A.J., Mirón-Gatón, J.M., Botella‐Cruz,
M., Pallarés, S., Sánchez-Fernández, D. 2022a. Thermal
tolerance and vulnerability to climate change in subterranean species: a case
study using an Iberian endemic pseudoscorpion. Insect Conservation and
Diversity 15: 181-190.
Colado, R., Pallarés, S., Fresneda, J., Mammola, S., Rizzo, V., Sánchez‐Fernández,
D. 2022b. Climatic stability, not average habitat temperature, determines
thermal tolerance of subterranean beetles. Ecology 103: e3629.
Culver, D.C.,
Pipan, T. (eds.) 2019. The biology of caves and
other subterranean habitats. 2nd edition. Oxford University
Press, Oxford, Reino Unido.
Danielopol, D.L., Griebler, C., Gunatilaka A., Notenboom, J. 2003. Present
state and future prospects for groundwater ecosystems. Environmental
Conservation 30: 104-130.
Darwin, C. 1859. On the origin of species. John Murray. Londres,
Reino Unido.
Escobar, L.E., Medina-Vogel,
G., Peterson, A.T. 2014. Potential for spread of the
white-nose fungus (Pseudogymnoascus destructans) in the Americas: use of
Maxent and NicheA to assure strict model transference. Geospatial
Health 9: 221-229.
Galán, C. 2010. Evolución de la
fauna cavernícola: mecanismos y procesos que explican el origen de las especies
troglobias. Boletín de la Sociedad Venezolana de Espeleología 44: 22.
Giordano, M. 2009. Global
groundwater? Issues and solutions. Annual Review of
Environment and Resources 34: 153-178.
Griebler,
C., Avramov, M. 2015. Groundwater ecosystem
services: A review. Freshwater Science 34: 3.
Grimm, V., Johnston, A.S., Thulke, H.H., Forbes, V.E., Thorbek, P. 2020.
Three questions to ask before using model outputs for decision support.
Nature Communications 11: 1-3.
Hadley, N.F., Ahearn, G.A., Howarth, F.G. 1981 Water and metabolic
relations of cave adapted and epigean lycosid spiders in Hawaii. Journal of
Arachnology 9: 215–222.
Howarth, F.G. 1980. The zoogeography of specialized cave animals: a
bioclimatic model. Evolution, 394-406.
Howarth,
F.G., Moldovan, O.T. 2018. The Ecological
Classification of Cave animals and their adaptations. En: Moldovan, O.T,
Kovác, L., Halse, S. (Eds.), Cave Ecology, pp 41-67. Springer, Nueva
York, Estados Unidos.
Iannella, M., Fiasca, B.,
Di Lorenzo, T., Biondi, M., Di Cicco, M., Galassi, D. M. 2020. Jumping into the grids: mapping biodiversity hotspots in groundwater
habitat types across Europe. Ecography 43: 1825-1841.
IPCC. 2022. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution
of Working Group II to the Sixth Assessment Report of the Intergovernmental
Panel on Climate Change. [H.-O. Pörtner, D.C. Roberts, M. Tignor, E.S.
Poloczanska, K. Mintenbeck, A. Alegría, M. Craig, S. Langsdorf, S. Löschke, V.
Möller, A. Okem, B. Rama (eds.)]. Cambridge University Press, Cambridge, UK and
New York, NY, Estados Unidos.
3056 pp. https://doi.org/10.1017/9781009325844
IPBES. 2019. Summary for policymakers of the global assessment report on
biodiversity and ecosystem services of the Intergovernmental Science-Policy
Platform on Biodiversity and Ecosystem Services. Diaz, S., Settele, J.,
Brondizio, E.S., Ngo, VH.T., Guèze, M., Agard, J., Arneth, A., Balvanera, P.,
Brauman K.A. Butchart, S.H.M et al. (Eds.), IPBES Secretariat. Bonn, Germany. 56
pages. https://doi.org/10.5281/zenodo.3553579
Jeannel, R. 1943. Les fossiles vivants des cavernes. Gallimard. París,
Francia.
Juan, C., Guzik, M.T., Jaume,
D., Cooper, S.J. 2010. Evolution in caves: Darwin’s ‘wrecks of
ancient life’ in the molecular era. Molecular Ecology 19:
3865–3880.
Jiménez‐Valverde, A., Lobo, J.M.,
Hortal, J. 2008. Not as good as they seem: the importance of
concepts in species distribution modelling. Diversity and Distributions
14: 885-890.
Kunz, T.H., Braun de Torrez, E., Bauer, D., Lobova, T., Fleming, T.H.
2011. Ecosystem services provided by bats. Annals of the New York Academy of
Sciences 1223: 1–38.
Mammola, S.,
Isaia, M. 2017. Rapid poleward distributional
shifts in the European cave‐dwelling Meta spiders under the influence of competition dynamics. Journal
of Biogeography 44(12): 2789-2797.
Mammola, S.,
Leroy, B. 2018. Applying species distribution
models to caves and other subterranean habitats. Ecography 41:
1194-1208.
Mammola, S., Cardoso, P., Culver, D.C., Deharveng, L., Ferreira, R.L.,
Fišer, C., Zagmajster, M., et al. 2019a. Scientists' warning on the
conservation of subterranean ecosystems. BioScience 69(8): 641-650.
Mammola, S., Piano, E., Cardoso, P., Vernon, P., Domínguez-Villar, D.,
Culver, D.C., et al. 2019b. Climate change going deep: The effects of global
climatic alterations on cave ecosystems. The Anthropocene Review 6:
98–116.
Mammola, S., Piano, E., Malard, F., Vernon, P., Isaia, M. 2019c. Extending
Janzen’s hypothesis to temperate regions: a test using subterranean ecosystems.
Functional Ecology 33: 1638-1650.
Mammola, S., Amorim, I.R., Bichuette, M.E., Borges, P.A., Cheeptham, N.,
Cooper, S.J., et al. 2020. Fundamental research questions in subterranean
biology. Biological Reviews 95: 1855-1872.
Mammola, S., Meierhofer, M.B., Borges, P.A., Colado, R., Culver, D.C.,
Deharveng, L. et al. 2022. Towards evidence-based conservation of subterranean
ecosystems. Biological Reviews 97: 1476-1510.
McGowan, N.E., Roche, N., Aughney, T., Flanagan, J., Nolan, P., Marnell,
F., Reid, N. 2021. Testing consistency of modelled predictions of the impact of
climate change on bats. Climate Change Ecology 2: 100011.
Nagy, J.A., Bartholy, J., Pongrácz, R., Pieczka, I., Breuer, H.,
Hufnagel, L. 2017. Analysis of the impacts of global warming on European bat
species’s range area in the 21st century using regional climate model
simulation. Quarterly Journal of the Hungarian Meteorological Service 121(3):
285-301.
Nicolosi, G., Mammola, S., Verbrugge, L., Isaia, M. 2023. Aliens in caves: the global dimension of biological invasions in
subterranean ecosystems. Biological Reviews 98(3), 849-867. https://doi.org/10.1111/brv.12933
Pallarés, S., Colado, R.,
Pérez‐Fernández, T.,
Wesener, T., Ribera, I., Sánchez-Fernández, D. 2019. Heat
tolerance and acclimation capacity in subterranean arthropods living under
common and stable thermal conditions. Ecology and Evolution 9:
13731-13739.
Pallarés, S., Sanchez-Hernandez, J.C., Colado, R., Balart-García, P., Comas,
J., Sánchez-Fernández, D. 2020. Beyond survival experiments: using biomarkers
of oxidative stress and neurotoxicity to assess vulnerability of subterranean
fauna to climate change. Conservation Physiology 8: coaa067.
Pallarés, S., Colado, R., Botella-Cruz, M., Montes, A., Balart-García, P.,
Bilton, D.T., et al. 2021. Loss of heat acclimation capacity could leave
subterranean specialists highly sensitive to climate change. Animal
Conservation 24: 482-490.
Peterson, A.T., Soberón, J., Pearson, R.G., Anderson, R.P., Martínez-Meyer,
E., Nakamura, M., et al. (eds.) 2011. Ecological Niches and Geographic
Distributions (MPB-49). Princeton University Press, Princenton, NJ, Estados
Unidos.
Raschmanová, N., Šustr, V., Kovacˇ, Ľ., Parimuchova, A., Devetter, M. 2018.
Testing the Climatic Variability Hypothesis in Edaphic and Subterranean
Collembola (Hexapoda). Journal of Thermal Biology 78: 391–400.
Razgour, O. 2015. Beyond species distribution modeling: a landscape
genetics approach to investigating range shifts under future climate
change. Ecological Informatics 30: 250-256.
Razgour, O., Juste, J., Ibáñez, C., Kiefer, A., Rebelo, H., Puechmaille, S.
J., Jones, G., et al. 2013. The shaping of genetic variation in edge‐of‐range
populations under past and future climate change. Ecology Letters
16(10): 1258-1266.
Razgour, O., Salicini, I.,
Ibáñez, C., Randi, E., Juste, J. 2015. Unravelling the
evolutionary history and future prospects of endemic species restricted to
former glacial refugia. Molecular Ecology 24(20): 5267-5283.
Razgour, O., Forester, B., Taggart, J.B., Bekaert, M., Juste, J., Ibáñez,
C., Manel, S., et al. 2019. Considering adaptive genetic variation in climate
change vulnerability assessment reduces species range loss projections. Proceedings
of the National Academy of Sciences 116(21): 10418-10423.
Reboleira, A.S., Borges, P.A.V., Gonçalves, F., Serrano, A., Oromí, P. 2011.
The subterranean fauna of a biodiversity hotspot region - Portugal: an overview
and its conservation. International Journal of Speleology 40: 23–37.
Ripple, W.J., Wolf, C., Newsome, T.M., Gregg, J.W., Lenton, T.M., Palomo,
I., et al. 2021. World scientists’ warning of a climate emergency 2021. BioScience
71: 894-898.
Rizzo, V., Sánchez-Fernández, D., Fresneda, J., Cieslak, A., Ribera, I.
2015. Lack of evolutionary adjustment to ambient temperature in highly
specialized cave beetles. BMC Evolutionary Biology 15: 1-9.
Román-Palacios,
C., Wiens J.J. 2020. Recent responses to climate
change reveal the drivers of species extinction and survival. Proceedings of
the National Academy of Sciences 117: 4211-4217.
Rosa, G.,
Penado, A. 2013. Rana iberica (Boulenger, 1879)
goes underground: subterranean habitat usage and new insights on natural
history. Subterranean Biology 11: 15-29.
Salgado, J.M., Sendra, A.,
Rodríguez, P. 2022. Listado de especies y subespecies cavernícolas (hipogeas)
consideradas como troglobias y estigobias de la península ibérica e islas
Baleares. Monografías electrónicas SEA, vol. 11 (30-VI-2022). Sociedad
Entomológica Aragonesa (S.E.A.).
Sánchez-Fernández,
D., Rizzo, V., Cieslak, A., Faille, A., Fresneda, J., Ribera, I. 2016. Thermal niche estimators and the capability of poor dispersal
species to cope with climate change. Scientific Reports 6(1): 1-8.
Sánchez-Fernández,
D., Rizzo, V., Bourdeau, C., Cieslak, A., Comas,
J., Faille, A. et al. 2018. The deep subterranean environment as a potential
model system in ecological, biogeographical and evolutionary research. Subterranean
Biology 25: 1-7.
Sánchez-Fernández,
D., Galassi, D.M.P., Wynne, J.J., Cardoso, P.,
Mammola, S. 2021. Don't forget subterranean ecosystems in climate change
agendas. Nature Climate Change 11: 458– 459.
Siebert, S., Burke, J., Faures, J.M., Frenken, K., Hoogeveen, J., Döll, P.,
et al. 2010. Groundwater use for irrigation–a global inventory. Hydrology
and Earth System Sciences 14: 1863-1880.
Tikhonov, G., Opedal, Ø.
H., Abrego, N., Lehikoinen, A., de Jonge, M. M., Oksanen, J., et al. 2020. Joint species distribution modelling with the R‐package
Hmsc. Methods in Ecology and Evolution 11: 442-447.
United
Nations 2019. UN Report: Nature’s Dangerous
Decline ‘Unprecedented’; Species Extinction Rates ‘Accelerating.’ Available
at: https://www.un.org/sustainabledevelopment/blog/2019/05/nature-decline-unprecedented-report/. [Accedido el 15 Abril 2022].
Vaccarelli, I., Colado,
R., Pallares, S., Galassi, D.M., Sanchez-Fernandez, D., Di Cicco, M., et al.
2023. A global meta-analysis reveals multilevel and
context-dependent effects of climate change on subterranean ecosystems. One
Earth 6: 1510-1522
Wynne, J.J., Howarth, F.G., Mammola, S., Ferreira, R.L., Cardoso, P.,
Lorenzo, T.D., et al. 2021. A conservation roadmap for the subterranean biome. Conservation
Letters 14: e12834.
Yoder, J.A., Benoit, J.B., LaCagnin, M.J., Hobbs, H.H. 2011. Increased
cave dwelling reduces the ability of cave crickets to resist dehydration. Journal
of Comparative Physiology B 181: 595-601.